Structural basis of Integrator-dependent RNA polymerase II termination
science-easyview
Integrator에 의존하는 RNA polymerase II termination의 구조적 기반
Abstract
Integrator 복합체는 유전자의 프로모터 근처 영역에서 RNA polymerase II (Pol II)를 termination할 수 있습니다. 이전 연구에서 Integrator가 Pol II, DRB sensitivity-inducing factor (DSIF) 및 negative elongation factor (NELF)로 구성된 중단된 염기화 복합체에 결합하고 새로운 RNA 전사체를 절단하는 방법에 대해 알려졌습니다. 그러나 Integrator가 Pol II를 DNA template에서 제거하는 방법은 아직 설명되지 않았습니다.
이번 연구에서는 서로 다른 기능적 상태에서 완전한 Integrator–PP2A 복합체의 세 가지 cryo-EM 구조를 제시합니다. pre-termination complex의 구조는 이전에 밝혀지지 않았던, 전구의 INTS10–INTS13–INTS14–INTS15 모듈을 보여줍니다. 이 모듈은 ‘독침 (sting)’을 사용하여 DSIF DNA 클램프를 열고 termination을 용이하게 할 수 있습니다. Termination 후 복합체의 구조는 이전에 밝혀지지 않았던 INTS3 subunit과 관련된 sensor of single-stranded DNA complex (SOSS) factor가 Pol II가 종료된 후 Integrator에 재결합하는 것을 방지하는 방법을 보여줍니다. 비활성화된 폐쇄된 구조의 자유 Integrator–PP2A 복합체 구조는 INTS6이 PP2A 인산화효소의 활성 부위를 차단한다는 것을 보여줍니다.
이러한 결과는 Integrator가 Pol II transcription을 termination하는 방법에 대한 모델을 제시하며, 이 과정은 주요한 재배치를 포함한 세 단계로 진행됩니다.
Figures
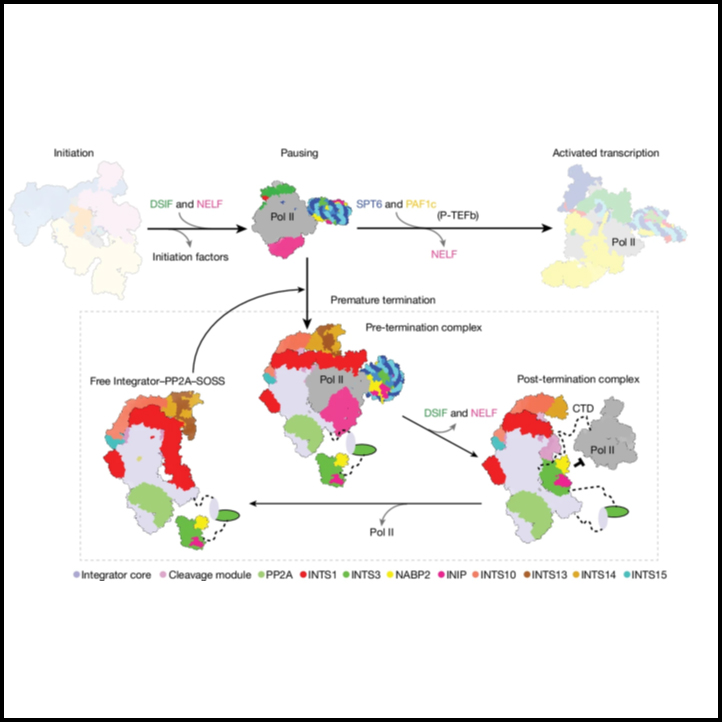
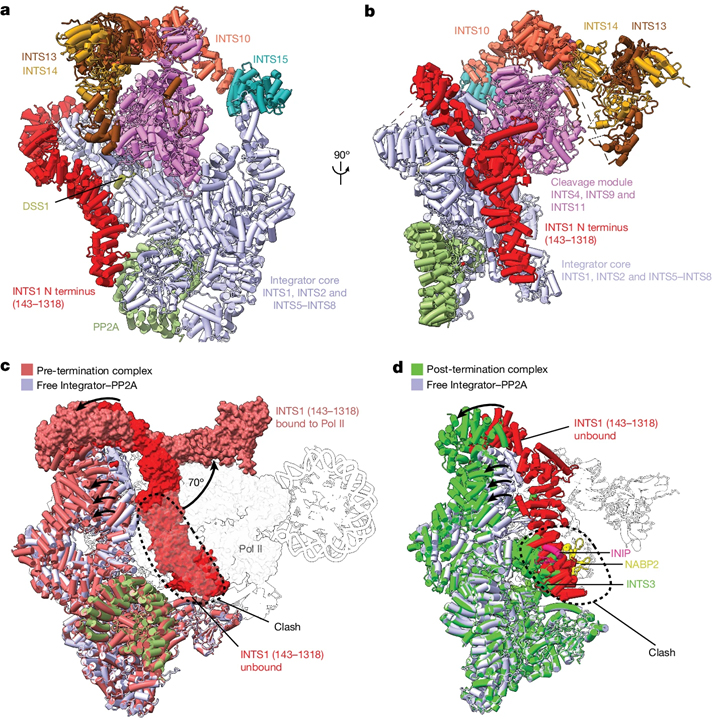
Structure of the pre-termination complex
[Figure 1] Pre-termination complex와 Integrator 활동의 cryo-EM 구조 (A) 복합체의 전체적인 구조. (B) 복합체의 대안적인 관점. (C) Integrator가 PEC에 결합된 RNA를 절단하고 분해하는 모습. (D) 시간에 따른 WT Integrator에 의한 PEC 결합 RNA의 분해.
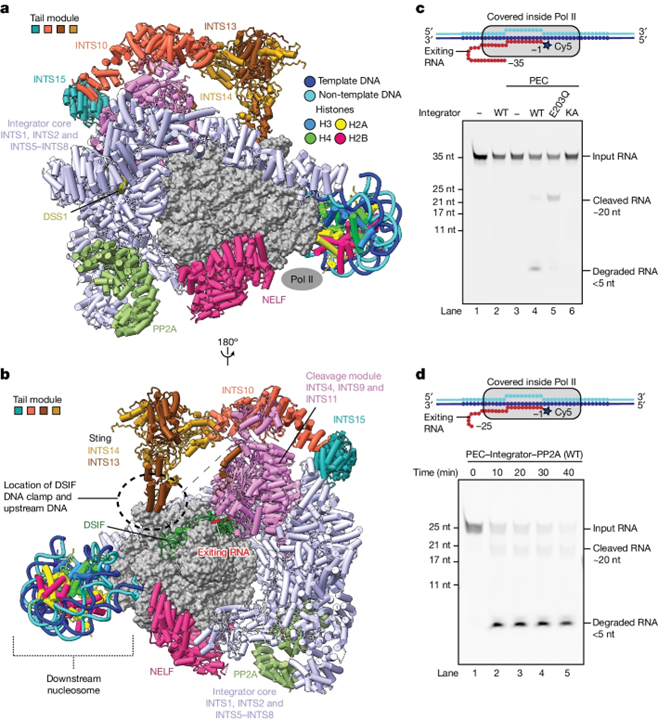
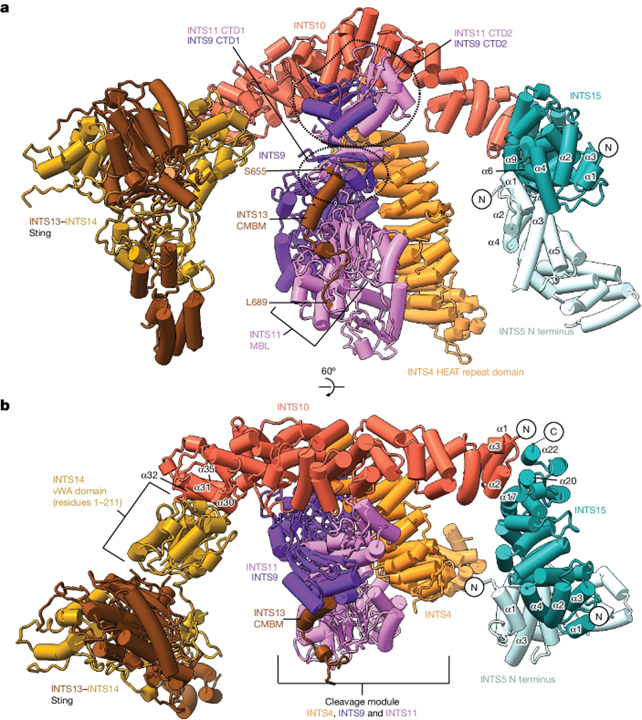
Structure of the Integrator tail module
[Figure 2] Integrator 꼬리 모듈의 구조 (A) 꼬리 모듈의 카툰 렌더링. (B) 대안적인 관점.
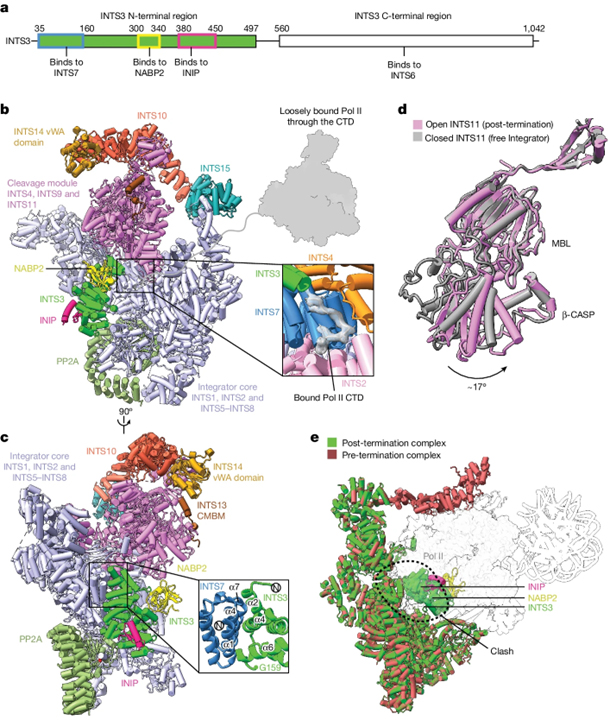
Structure of the post-termination complex
[Figure 3] post-termination complex의 cryo-EM 구조. (A) Integrator 하위단위 INTS3의 조직. (B) 종료 후 복합체의 cryo-EM 구조. (C) 복합체의 대안적인 관점. (D) 종료 구조에서의 열린 형태 구조. (E) pre-termination complex와 post-termination complex의 구조.
Complete Integrator–PP2A structure
[Figure 4] 자유 Integrator-PP2A 복합체의 cryo-EM 구조 (A) 자유 Integrator-PP2A 복합체의 cryo-EM 구조. (B) 복합체의 대안적인 관점. (C) pre-termination complex와 자유 Integrator-PP2A 복합체의 구조. (D) 자유 Integrator-PP2A 복합체와 post-terminatino complex의 구조.
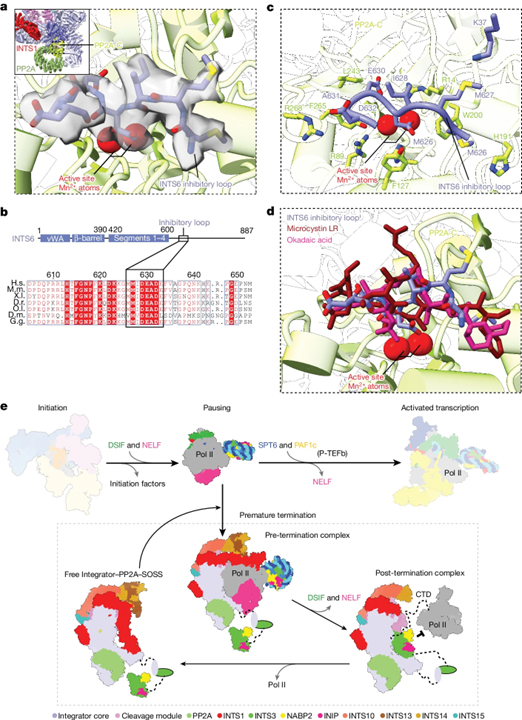
INTS6 blocks the PP2A active site
[Figure 5] Integrator 하위단위 INTS6은 PP2A를 조절하며, Integrator 의존적 Pol II 종료 주기의 모델 (A) 자유 Integrator-PP2A 구조에서 PP2A-C 활성부 앞에 결합된 INTS6 억제 루프의 cryo-EM 밀도를 보여주는 확대된 보기. (B) 상단, Integrator 하위단위 INTS6의 조직, 비정렬된 C-term에 있는 억제 루프의 위치.하단, 종 다양성에서 억제 루프의 보전을 보여주는 단백질 서열 정렬. (C) INTS6 억제 루프 및 PP2A-C 상호작용의 세부 내용을 보여주는 확대된 보기. (D) INTS6 억제 루프와 PP2A-C 억제제 okadaic acid 및 microcystin-LR의 비교. (E) Integrator 의존적 Pol II 종료에 대한 모델.
Disscussion
Integrator에 의한 조기 종료는 Pol II 전사를 조절하는 중요한 방법으로 부상하고 있지만, Integrator가 Pol II를 종료하는 방법에 대한 이해는 제한적입니다. 여기에서는 세 가지의 완전한 Integrator의 cryo-EM구조를 제시합니다. 하나는 종료 전 상태의 PEC에 결합된 것이고, 다른 하나는 종료 후 상태의 Pol II에 결합된 것이며, 나머지 하나는 비활성 상태의 완전한 자유 복합체의 구조입니다. pre-termination complex의 구조는 이전에 해결되지 않았던 Integrator 꼬리 모듈을 보여주며, 이는 전갈의 꼬리 모양으로, 쐐기를 포함합니다. 꼬리 모듈은 쐐기를 위치시키고 DSIF DNA clamp를 이동시키고 상류 DNA와 간섭하여 Integrator 코어와 절단 모듈에 결합합니다. 종료 후 구조는 INTS3 N-term이 Integrator의 개방된 구조에 있는 INTS7에 결합하고 종료 후에 Pol II가 Integrator에 재결합하는 것을 방지합니다. 마지막으로, 자유 Integrator-PP2A 복합체의 구조는 PEC가 없는 경우, 복합체가 비활성 상태로 취할 수 있으며, INTS1 N-term이 INTS3와의 결합을 방해하고 INTS6이 PP2A-C 활성 부위를 차지합니다.
생화학적 데이터와 이전 연구와 함께, 우리의 구조는 Integrator-의존적 Pol II 종료를 위한 세 단계 모델을 제안합니다. 먼저, 비활성 Integrator-PP2A 복합체가 PEC에 결합하기 위해 구조적 재배열을 겪어 PP2A 인산화 효소를 포지셔닝하고, Pol II CTD의 인산화를 가능하게 함으로써 P-TEFb의 kinase 활동에 반항합니다. 둘째, Integrator 꼬리 모듈은 DSIF DNA clamp와 upstream DNA를 이동시키고, Integrator endonuclease INTS11이 나선 상이였던 nascent RNA를 절단하고 이를 추가로 분해하여 DNA-RNA 하이브리드를 풀어내고 전사 버블을 붕괴시킵니다. 이로써 Pol II가 DNA로부터 해방됩니다. 우리는 RNA 분해와 DNA 해방이 연결되어 있을 것으로 추정합니다. 왜냐하면 꼬리와 절단 모듈은 INTS13 CMBM에 의해 연결되어 있습니다. 세 번째로, Pol II가 Integrator로부터 해방되며 INTS3와 관련된 SOSS 요인들에 의해 Pol II와의 복합체 형성이 스테릭적으로 방지됩니다. 마지막으로, Integrator는 닫힌, 비활성 구조로 변화하여 또 다른 종료 주기에 준비됩니다.
Integrator-의존적 Pol II 종료를 위한 이 모델은 다른 인자들에 의한 Pol II 종료의 현재 모델과 개념적으로 유사합니다. 특히, 단백질 코딩 유전자 끝에서 적용되는 torpedo 모델은 5′ – 3′ exonuclease XRN2를 포함하며, 이는 5′ 말단에서 nascent RNA를 분해하고 DNA-RNA 하이브리드를 풀어내어 전사 버블을 붕괴시키고 Pol II 종료를 유발합니다. Integrator endonuclease INTS11은 종속성이 있는데, 이는 복제 종속적 histone 종료를 위한 torpedo nuclease로 제안되었습니다. Integrator-의존적 프로모터-근접 종료와 비코딩 Pol II 전사의 종료는 Pol II의 NELF-의존적 인터럽트가 필요하므로, INTS11의 추정된 5′-3′ exonuclease 활동은 Pol II에 의해 결합된 DNA-RNA 하이브리드를 풀어낼 수 있을 것으로 생각됩니다. INTS11이 첫 번째 절단 이후의 Pol II 내부에 있는 RNA에 도달하는 방법은 불명확합니다. INTS11이 첫 번째 절단 이후 Pol II와 연관된 RNA의 5′ 끝에 여전히 결합되어 있을 것으로 추측됩니다. 왜냐하면 INTS11 활성 부위 홈은 절단 부위 이전에 나타난 나선 RNA의 약 5nt를 수용할 수 있기 때문입니다. 이로써 Integrator는 RNA의 5′ 끝을 분해하면서 RNA 3′ 끝으로 이동하고 이 과정에서 Pol II를 전진시킬 수 있습니다. 우리의 모델과 일관되게, 절단기-의존적 metazoans 동물에서의 Pol II 종료는 XRN2에 독립적입니다. 그러나 Integrator만으로 종료가 이루어지는 경우와 Integrator가 종료를 달성하기 위해 XRN2와 협력할 때에는 언제인지 아직 알려지지 않았습니다.