
Arabidopsis의 내열성에 필요한 온도 센서 TWA1

Abstract
과도한 온도에 노출된 식물은 생리적 도전에 대처하고 장기적인 적응을 촉진하기 위해 열 스트레스 반응을 활성화합니다. 그러나 세포 온도를 감지하여 내열성을 유도하는 메커니즘은 아직 명확하지 않습니다.
이 연구에서는 TWA1이 기본적이고 획득된 내열성에 필요한 온도 감지 전사 조절 인자임을 보여줍니다. 높은 온도에서 TWA1은 그 구조를 변화시키며, 억제자 복합체 형성을 위해 JASMONATE-ASSOCIATED MYC-LIKE (JAM) 전사 인자, TOPLESS (TPL), TOPLESS-RELATED (TPR) 단백질과 물리적으로 상호작용할 수 있게 합니다. TWA1은 아미노 말단의 매우 가변적인 영역을 통해 기능하는 중요한 온도 감지 역할을 하는 본질적으로는 무질서한 단백질로 예측됩니다. 높은 온도에서 TWA1은 nuclear subdomain에 축적되며, JAM2 및 TPL과의 물리적 상호작용은 이 nuclear subdomain에 제한되는 것으로 보입니다. Heat shock transcription factor A2 (HSFA2)와 heat shock protein의 전사적 상향 조절은 TWA1에 의존하며, TWA1 상동체는 열 스트레스의 초기 신호 전달에서 센서 기능과 일치하는 다양한 온도 임계값을 제공합니다.
식물 온도 센서의 식별은 육종 및 생명공학을 통해 작물의 열 적응 반응을 조정할 수 있는 분자적 도구를 제공하며, 열유전학을 위한 민감한 온도 스위치를 제공합니다.
Figures
Identification of TWA1

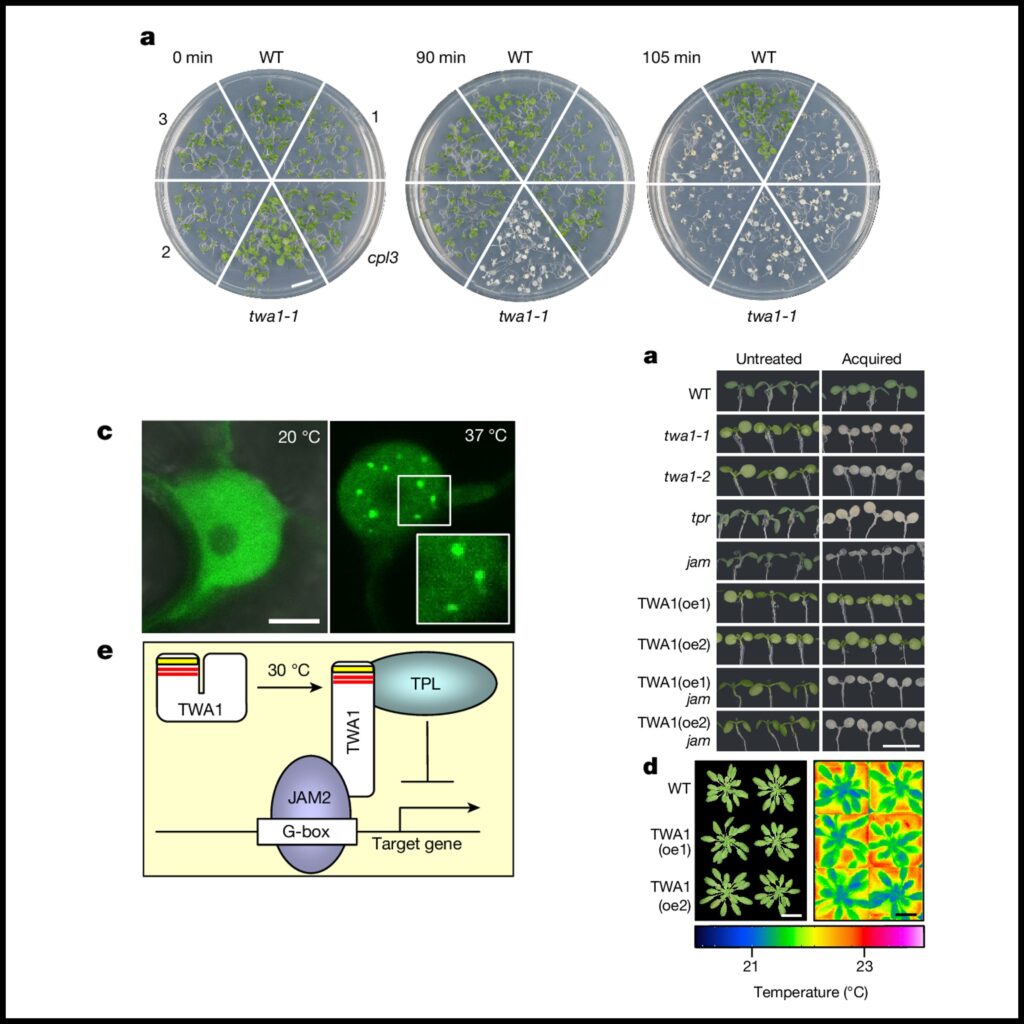
[Figure 1] TWA1 유전자 자리
(A) WT A. thaliana와 twa1-1 돌연변이를 45°C에서 90분 또는 105분 동안 열 스트레스 를 주고 5일 후에 찍은 이미지.
(B) WT, twa1-1, twa1-2 식물의 획득된 내열성, 그리고 TWA1 유전자로 보완된 돌연변이 식물의 열 노출 후 5일 이내의 뿌리 길이 평가 결과.
(C) 5일 된 묘목에서 17°C 또는 27°C에서 16시간 동안 1µM ABA에 노출된 경우의 ABA-유도성 pHB6:LUC 발현.
왼쪽, 리포터를 발현하는 WT와 twa1-1 식물의 사진과 빛 방출. 오른쪽, 묘목 당 relative light units으로 제공된 리포터 활성.
(D) 최대 점수 1을 가진 본질적으로 무질서한 영역의 예측과 TWA1의 도식.
상단, Highly variable region (HVR, 노란색) 및 두 개의 EAR 도메인 (빨간색) . 돌연변이체에서 TWA1의 파괴는 화살표로 표시. 하단, A. thaliana (A.t.)와 A. lyrate (A.l.)의 HVR의 아미노산 상동 (빨간색).
Thermosensor function

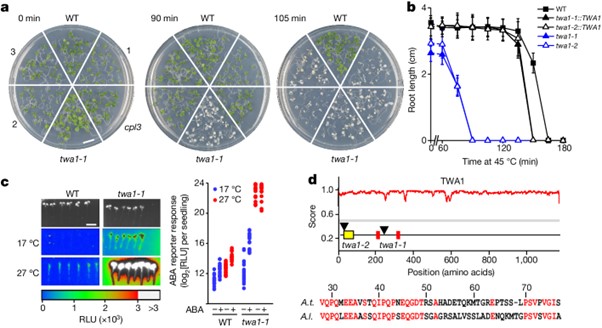
[Figure 2] TWA1의 온도 센서 기능
(A-C) TWA1 및 TWA1 변형체는 잎 원형질체에서 ABA-유도성 pRD29B:LUC 발현 억제.
(A) WT, twa1-1, twa1-2 식물의 원형질체를 TWA1 또는 twa1-1 발현 카세트로 형질전환 하고 10µM ABA와 함께 25°C에서 16시간 동안 배양 후 측정한 RLU 결과.
(B) 다양한 ABA 양에 따른 TWA1 작용.
(C) 10^5 twa1-2 원형질체당 0.3µg의 effector DNA로 다양한 배양 온도에서의 TWA1 및 변형체의 작용.
변형체: TWA1(ΔN), A. lyrata AlTWA1, SaTWA1, TWA1(AlHVR)
(D) 상단, 효모 핵 내에서 GFP-태그가 된 TWA1 및 twa1 산물과 온도 의존적 phase separation. 하단, 22°C에서 TWA1(AlHVR)의 nuclear subdomain 형성.
(E) 분자 내 FRET–FLIM 분석으로 밝혀진 TWA1 아미노- 및 카복시-말단 도메인의 온도 유도 변화.
상단, 효모 핵 내 GFP FL의 mCherry–TWA1–GFP (TWA1)의 false-colour image. 하단, TWA1과 mCherry–GFP의 GFP FL.
(F,G) TWA1 및 변형체, 포함 twa1 산물과 JAM2 orthologues 결합에 대한 온도 의존성 Y2H 성장 분석.
TWA1-mediated transcriptional regulation

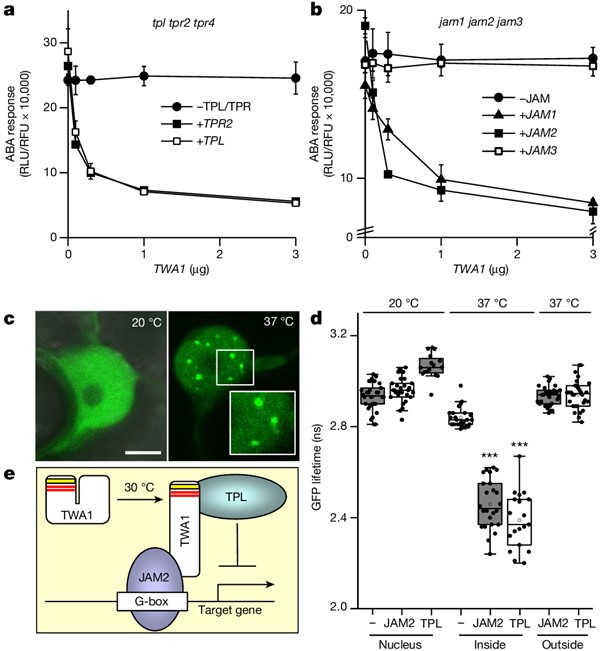
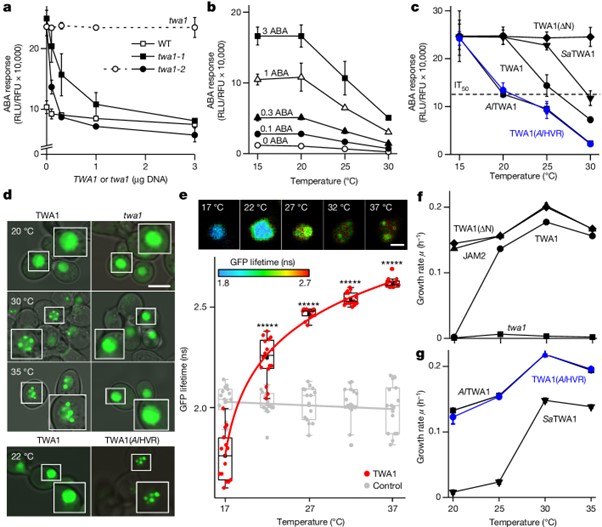
[Figure 3] TWA1 매개 전사 억제의 분자 메커니즘
(A) TPL과 TPR 공동억제자는 TWA1 억제 리포터 발현에 필요함을 보이는 RLU 결과.
tpl tpr2 tpr4 삼중 돌연변이 식물의 원형질체를 TWA1, TPL, TPR2 effector DNA로 형질전환.
(B) JAM1 및 JAM2 co-expression은 jam1 jam2 jam3 식물의 원형질체에서 TWA1 반응 복구.
(C,D) N. benthamiana 표피 세포의 nuclear subdomain에서 JAM2 및 TPL에 대한 GFP–TWA1의 축적 및 결합.
(C) 핵에서의 GFP 이미지.
(D) FRET–FLIM 분석.
(E) 억제 복합체 형성의 단순화된 모델.
[Fig. 3E] 온도가 20 °C에서 30 °C로 전환되면 TWA1의 구조적 변화가 유도되어 JAM2가 TWA1의 카복시 말단 부분과 결합하고 TPL이 EAR 모티프 (빨간색)를 통해 아미노 말단 도메인에 결합할 수 있게 됩니다. HVR (노란색)은 온도 감지에 필수적입니다. JAM2는 dimer로서 G-box 관련 cis-elements와 상호작용합니다. TPL은 tetramer이며 mediator complex의 subunit을 표적으로 합니다.
Thermotolerance conferred by TWA1

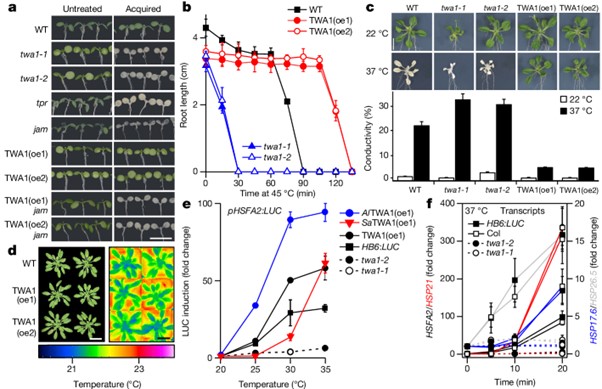
[Figure 4] TWA1에 의한 열 내성 및 heat shock 전사체 조절
(A) 각 조건에 따른 열 스트레스가 없거나 45 °C, 150분의 열 스트레스 후 1일 뒤에 촬영한 이미지.
(B) 5일 된 묘목의 기초 열 내성에 대한 뿌리 길이 평가 결과.
(C) 3주 된 식물을 37 °C에 24시간 노출하거나 22 °C에 유지한 후의 이미지와 이온 누출 결과.
(D) 22 °C에서 단일 낮 조건으로 38 일 동안 자란 식물의 사진과 false-colour로 된 열 이미지.
(E) 다양한 라인의 잎 원형질체에서 LUC 발현을 유도하는 HSFA2 프로모터의 온도 및 TWA1 의존적 유도.
(F) 20 °C에서 자라며 최대 20분 동안 37 °C에 노출된 5일 된 WT 및 twa1 묘목의 heat shock 전사체 변화.
[Fig. 4A] 획득된 열 내성은 TPL/TPR형 공동억제자와 JAM 전사인자에 의존하며, jam1 jam2 jam3 (jam) 식물에서 TWA1 발현(TWA1(oe) 라인)에 의해 부여된 열 내성 상실을 포함합니다. TWA1은 twa1-2 식물에서 바이러스 35S 프로모터 하에 발현되었습니다. tpr은 tpl tpr2 tpr4 유전형을 나타냅니다.
[Fig. 4D] TWA1(oe) 라인의 개선된 열 내성은 성장의 저하나 명백한 광합성 매개 변수의 변화 없이 나타납니다.
Disscussion
애기장대에서 TWA1의 열 감지 기능이 열 내성과 HSFA2 및 HSP 전사체의 적시 유도에 필요함을 밝혀냈습니다. HSR은 주요 HSF1 유형 TF의 활성화에 의해 촉발됩니다. 애기장대에서는 세 관련 HSFA1들이 중복적으로 HSR을 시작합니다. 이 마스터 TF 중 두 개의 비활성화는 여전히 HSFA2 유도와 열로 인한 이온 누출에 대한 내성을 허용합니다. 두 반응 모두 TWA1 결핍 식물에서는 손상되며, 이는 열 감지기가 HSFA1들보다 상위에 있거나 대안적으로 평행하게 작용함을 시사합니다. 지속적으로 활성화된 HSFA1 TF와 TWA1의 이소성 발현은 향상된 열 내성을 제공합니다. HSFA1에 의한 지속적인 열 내성은 사전 유도된 HSR 및 성장 패널티와 관련이 있으며, 이는 또한 HSFA2 및 HSFA3의 이소성 발현에 의해 나타납니다. 그러나 TWA1 매개 열 내성에서는 이러한 불리한 생리적 효과가 관찰되지 않았습니다. 또한 몇 분 내에 HSR 전사체의 유도는 기능적인 TWA1을 필요로 했습니다. 이 발견은 온도 상승에 대한 초기 신호 전달에서 TWA1의 유도 가능한 보호 기능을 지적합니다.
TWA1의 열 활성화는 HVR과 JAM2 및 억제 단백질의 결합을 조절하는 온도 유도 도메인 재배치를 포함합니다. TWA1의 역할은 열 감지기 EARLY FLOWERING 3 (ELF3)와 유사점을 가집니다. ELF3는 저녁 복합체의 구성 요소로서 애기장대의 생체 시계 및 개화 시기를 조절하는 전사 공동 억제기로 기능합니다. 열적으로 TWA1은 높은 온도에서 유전자 발현을 억제하고 온도 변화에 더 민감하게 반응하여 ELF3와 반대 방식으로 작용합니다. TWA1의 반응성은 Y2H 분석에서 성장률로부터 유도된 10°C 차이가 활동을 150배 변화시킬 수 있는 Q10 값 약 150에 해당합니다. 이 TWA1의 반응성은 뱀과 모기의 TRPA1 Ca2+ 채널 구성원을 포함하여 가장 열에 민감한 단백질들 중 하나인 것으로 알려져 있습니다. 진핵생물에서는 열 스트레스가 세포질 및 소기관 스트레스 과립을 유도하고, 이는 식물에서 선구적인 역할을 하며, 열 도전에 대한 세포 적응을 위해 단백질과 RNA의 가역적인 집합체인 nuclear subdomain 형성을 유도합니다. Liquid–liquid phase separation (LLPS)는 활성 유전자 발현 부위로서 다층 nuclear subdomain 형성에 중요한 역할을 합니다. 열 활성화 후 TWA1은 LLPS를 연상시키는 nuclear subdomain에 축적됩니다. Subdomain은 JAM2 및 TPL과의 TWA1 상호 작용 부위로, 가정된 LLPS에서 활성 억제 복합체의 형성을 나타냅니다. 본질적으로 무질서한 영역을 가진 단백질은 LLPS에 대한 경향이 있으며, 프리온 같은 도메인을 가진 식물 단백질을 포함합니다. 열 스트레스 하에서 효모 HSF1의 본질적으로 무질서한 영역은 응집체 형성과 단일 LLPS 내 전사를 위한 다중 염색질 상호 작용을 촉진합니다. 효모 및 포유류에서 HSF1 활성화로 이어지는 온도 감지 메커니즘은 아직 명확하지 않지만, 샤페론 HSF1 적정 모델에 대한 증거가 점점 늘어나고 있습니다.
TWA1 유형 센서는 식물에만 독특할 수 있습니다. TWA1은 20°C에서 30°C 사이의 아열 온도 상승을 감지하고 초기 HSR 전사체의 유도에 필요합니다. 그러나 4시간의 준비 단계 후, HSP 전사체 수준은 WT와 twa1 돌연변이체 사이에서 비슷하여 추가적인 전사체 상향 조절 모드를 암시합니다. 식물에서는 열에 의해 유도된 과산화수소 및 질소 산화물 증가가 HSFA1을 활성화하고 HSR에 적색 신호 통합을 허용합니다. TWA1은 또 다른 신호 통합 노드를 제공할 수 있습니다. 식물은 종종 단일 스트레스 반응을 통합하는 도전에 직면한 복합 스트레스를 경험합니다. 예를 들어, 수분 부족 하에서 ABA 매개 프롤린 증가는 보호성 삼투체를 제공하지만, 높은 프롤린 수준은 열 하에서 독성이 있습니다. TWA1은 아마도 온도를 ABA 및 JA 신호 전달과 통합하여 비생물적 및 생물적 신호를 각각 전달함으로써 적응 반응을 조율할 것입니다. 미래에는 TWA1 유형 온도 센서를 다른 열 특성으로 육종 및 생명공학을 통해 사용하여 작물의 적응 반응을 온난화 기후에 맞출 수 있을 것입니다. 또한 TWA1과 그 상동체는 열유전학의 신흥 분야에서 온도 스위치를 설계하기 위한 원형을 제공합니다.