
면역 미세 환경이 장 내 Treg 기능을 조절한다.

Abstract
장의 면역 시스템은 공생 미생물과 자가 항원에 대한 내성을 유지하면서 침입하는 병원체로부터 방어할 수 있도록 매우 적응되어 있다. 장의 복잡한 면역 환경에서 현지 세포들의 다양한 네트워크가 홈에오스타시스를 어떻게 확립하고 유지하는지를 이해하는 것은 염증성 질환과 같은 기능 이상이 발생한 후 내성을 다시 확립하는 방법을 이해하는 데 중요하다. T 조절(레귤러토리, Treg) 세포의 발달과 기능을 제어하는 세포 및 분자 상호작용은 확인되었지만, 미생물에 반응하는 Treg 세포의 기능을 형성하는 세포적 환경과 공간적 구획화에 대해서는 잘 알려지지 않았다. 여기에서 우리는 생체 내 실시간 영상, 광활성화 유도 단일 세포 RNA 시퀀싱, 공간 전사체 분석을 사용하여 내성 및 염증 설정에서 Helicobacter hepaticus에 반응하는 T 세포의 자연사를 공간과 시간을 통해 추적했다. 항원 자극은 조직 어디에서나 일어날 수 있지만, 내장 림프 조직에 포함되지 않은 라미나 프로피아가 효과적인 Treg (eTreg) 세포 기능을 지원하는 핵심 미세 환경이다. eTreg 세포는 그들의 미세 환경이 확립되면 안정적이지만, 염증이 촉발되면 구획화가 붕괴되어 라미나 프로피아에서 CD103+SIRPα+ 수지상 세포의 지배로 이어진다. 우리는 라미나 프로피아에서 CD206+ 대식세포와 eTreg 세포 사이의 추정 내성 유도 상호작용을 확인하고 이 상호작용을 지배할 가능성이 있는 수용체-리간드 쌍을 확인했다. 우리의 결과는 라미나 프로피아에서 내성의 공간적 메커니즘을 밝히고 현지 상호작용에 대한 지식이 내성 유도 치료의 차세대에 어떻게 기여할 수 있는지를 보여준다.
Figures
eTreg cells function in intestinal tissue
– eTreg cell의 장 조직에서 기능

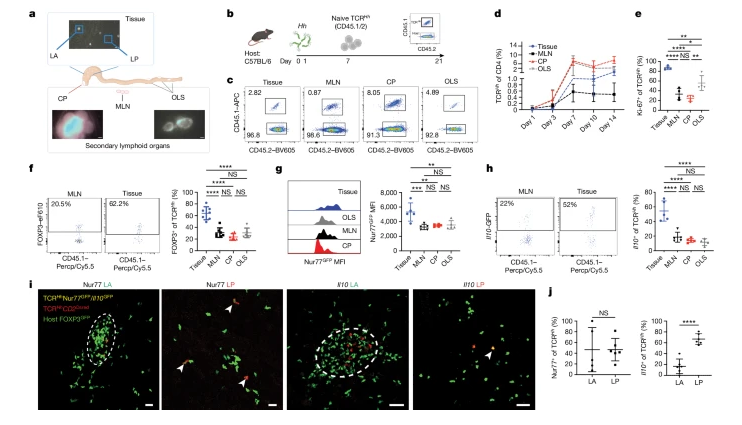
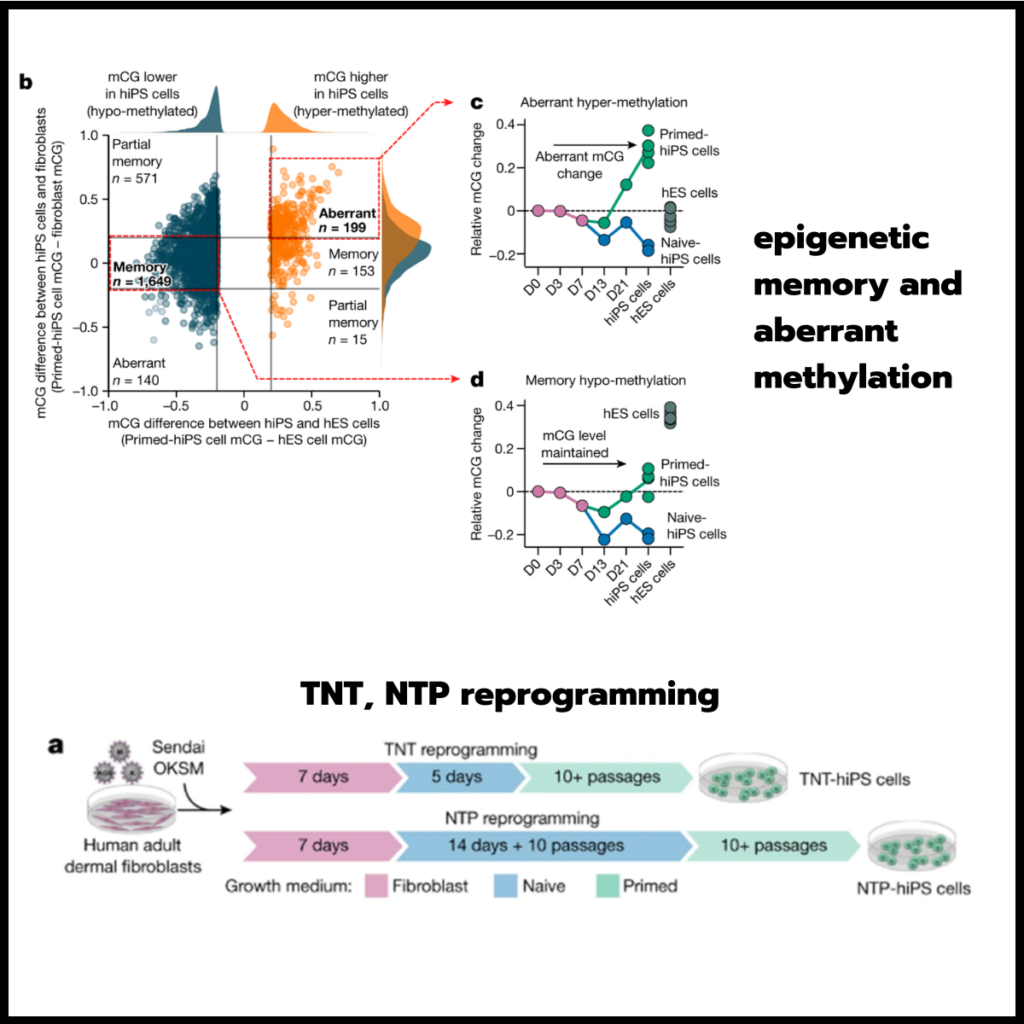
Fig. 1. LP 니치는 Hh 콜로니제이션에 대한 반응으로 IL-10 생성이 가장 높다.
(A) Hh에 대한 적응 반응이 발생하는 곳을 매핑하기 위해 야생형 마우스의 장 림프 조직 구획과 비림프 조직 구획을 특성화했습니다. 림프 조직은 MLN, 맹장 패치(CP) 및 원위 결장 조직 림프 구조(OLS)를 포함한 2차 림프 기관으로 구성되었습니다
(B,C,D) 나이브 TCRHhT 세포는 Hh-집락화된 숙주로 전달되었고(그림 1b 및 확장 데이터 그림 1d), 24시간 이내에 2차 림프 기관으로, 이후에는 약 1주일 후에 조직으로 전달되었습니다
(E) 림프구 구획에서 초기 활성화를 나타냄에도 불구하고(Extended Data Fig. 2a), 이동 후 14일째에 조직 내 TCRHhT 세포는 림프구에 비해 더 증식성이 높습니다
(F) 2차 림프 기관보다 조직에서 FOXP3+ Treg 세포로 분화된 TCRHhT 세포의 비율이 더 높았습니다
(G) 우리는 TCRHh 마우스를 Nur77GFP 마우스19(TCRHhNur77GFP)로 교배하고 이 마우스의 T 세포를 Hh-집락화된 숙주로 옮겼습니다. 전체 이동 집단에서 GFP 및 GFP 기하평균 형광 강도(MFI)를 발현하는 세포의 비율(TCR 신호 전달 측정)은 이동 후 11일째에 조직에서 가장 높았습니다.
(H) C57BL/6 Foxp3CD2Il10GFP 리포터 마우스20(TCRHhIl10GFP)에 교차된 T 세포를 Hh-집락화된 숙주로 이동시킨 결과, 이동 후 11일째에 Il10+TCRHhT 세포의 비율이 가장 높은 부위로 조직이 밝혀졌습니다
(I) 조직 내에서 TCRHhNur77GFP+ T 세포는 LA 내에 존재했습니다(그림 1i). TCRHhNur77GFP+ 세포는 또한 LA로부터 공간적으로 멀리 떨어진 LP 내에 위치했습니다
(J) 최근에 활성화된 T 세포의 이러한 고른 분포(그림 1j)는 TCR-주요 조직적합성 복합체 그룹 II(MHCII) 상호작용이 TCRHh T 세포와 LP-거주 항원 제시 세포(APC) 사이에서 발생하고/또는 TCR-자극된 TCRHh T 세포가 LA에서 LP로 빠르게 이동함을 시사합니다.
[Fig 1A] CD2DsRed 마우스에서 CP, 조직화된 림프 구조 및 LA의 대표적인 이미지를 포함하는 마우스 내장의 조직 및 림프 구조 다이어그램. 눈금 막대, 1,000 μm
[Fig 1B] 순진한 TCRHhT 세포를 Hh-집락화된 숙주로 이동시키는 도식
[Fig 1C] 나이브 TCRHHT 세포의 이동 후 14일 후 표시된 부위에서 TCRHT 세포의 대표적인 형광 활성화 세포 분류(FACS) 플롯
[Fig 1D] 순진한 TCRHhT 세포의 이동 후 표시된 시점에서 총 CD4 T 세포 중 TCRHh 세포의 빈도
[Fig 1E] 나이브 TCRHhT 세포의 이동 후 14일 후 표시된 부위에서 Ki-67+ TCRHh 세포의 빈도
[Fig 1F] 대표 FACS 플롯(왼쪽)과 나이브 TCRHh T 세포의 이동 후 14일째 TCRHh(오른쪽)의 Treg 세포의 빈도
[Fig 1G] 나이브 TCRHHhT 세포 이송 후 11일 후 TCRHhNur77GFPT 세포의 대표 히스토그램 오버레이(왼쪽) 및 GFP MFI
[Fig 1H] 대표 FACS 플롯(왼쪽) 및 나이브 TCRHHT 세포 이송 11일 후 TCRHHT 세포 중 Il10+ 세포의 빈도(오른쪽)
[Fig 1I] LA 및 LP에서 TCRHHnNur77GFP(좌측 및 중앙 좌측) 및 TCRHHIl10GFP(우측 및 중앙 우측) T세포의 대표적인 이미지. TCRHhNur77GFP 및 Il10-GFP 양성 세포는 흰색 화살촉으로 표시됩니다. 눈금 막대, 50 μm
[Fig 1J] LA 및 LP의 TCRHHT 세포 중 Nur77GFP(왼쪽) 및 Il10-GFP 양성(오른쪽) 세포의 빈도
Spatially and phenotypically distinct Treg cells
– 공간적으로 및 형질적으로 구별된 Treg 세포

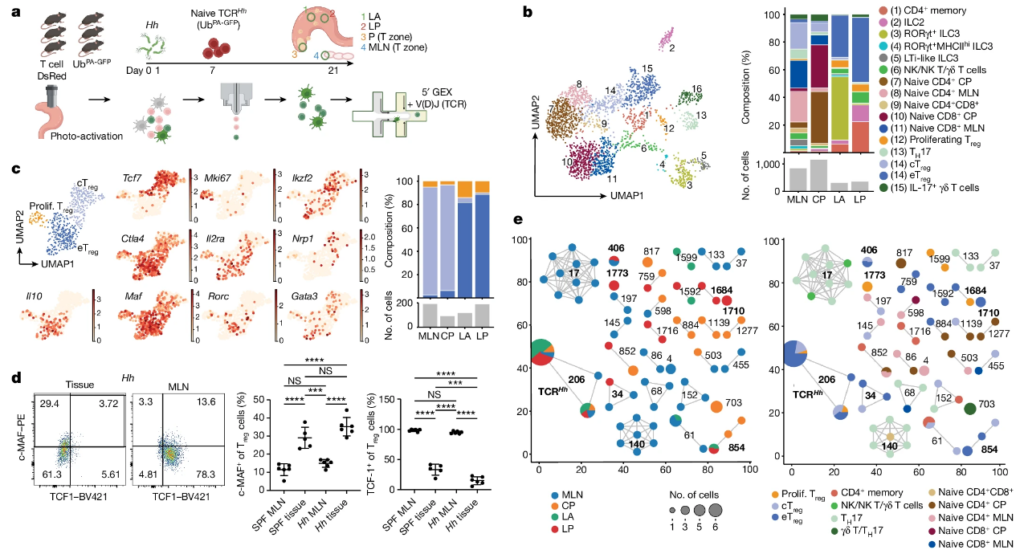
Fig. 2. cTreg 세포와 eTreg 세포 집단은 전사적으로 그리고 공간적으로 구별됩니다.
(A) TCRHHCD2DsRed 마우스에서 광활성화 가능한 GFP6(TCRHCD2DsRedUbPA-GFP)를 편재적으로 발현하는 마우스로 교차된 순진한 T 세포는 Hh-집락화된 CD2DsRedUbPA-GFP 호스트로 전달되어 호스트 세포와 기증자 세포가 모두 광활성화될 수 있었습니다. DsRed는 기증자 세포를 시각화하고 호스트 림프 조직의 T 세포 영역을 표시하는 데 사용되었습니다. 우리는 2광자 현미경을 사용하여 4개 영역(MLN 및 CP, LA 및 LP의 T 세포 영역)에서 광활성화된 GFP로 기증자와 호스트 세포를 변환했습니다
(B) T 세포 및 ILC 구획은 전사적으로 구별되는 16개의 하위 집합으로 구성됩니다
(C) Treg 세포의 하위 군집링은 2차 림프 기관과 조직 사이에 차등적으로 분포된 3개의 하위 집단을 나타냈습니다
(D) Treg 세포의 편향되지 않은 클러스터링은 cTreg-eTreg 분열을 기반으로 세포를 분리하기 때문에, 이는 이러한 발달 상태가 이전에 설명한 존재론적 구별보다 지배적인 마이크로치 내의 상호 작용을 기반으로 최우선적인 환경 각인을 나타냄을 시사합니다. cTreg 대 eTreg 표현형은 단백질 수준의 특정 병원체 자유 (SPF) 마우스에서도 관찰되며, 이는 이러한 뚜렷한 Treg 세포 표현형이 Hh 설정에 국한되지 않음을 시사합니다
(E) 숙주 세포와 전송된 세포가 다운스트림 분석과 비교 가능한지 여부를 결정하기 위해 T 세포 클론형 분석을 수행했습니다. 우리는 TCRHhT 세포(클론형 206)를 검출했는데, 이는 이 세포들이 조직에서 eTreg 세포가 되고 모든 조직에서 가장 확장된 클론을 나타낸다는 것을 보여주었습니다
[Fig 2A] 광활성화, 세포 분류 및 scRNA-seq. DC, 수지상 세포; 림프, 림프; Mac, 대식세포; 모노, 단핵구를 갖는 6개의 CD2DsRedUbPA-GFP 숙주로의 TCRHCD2DsRedUbPA-GFP 세포의 전달을 개략적으로 설명하는 것.
[Fig 2B] 왼쪽, 모든 위치에 걸친 T 및 ILC 부분집합의 UMAP 시각화 오른쪽, 림프 부분집합의 분포(위) 및 세포 수(아래). NK, 자연살해자.
[Fig 2C] 모든 위치(왼쪽 위)에 걸친 Treg 세포 부분 집합의 UMAP 시각화 및 선택된 유전자에 대한 발현 데이터의 UMAP 플롯에 오버레이(중간; 색상은 상대적 발현을 나타냄). 오른쪽, Treg 세포 부분 집합(위) 및 세포 번호(아래) 분포.
[Fig 2D] 좌측, 조직 내 Treg 세포 및 Hh-집락화 마우스의 MLN에 대한 c-MAF 대 TCF1의 대표 FACS 플롯. 조직 내 c-MAF+ (중심) 및 TCF1+ (우측) Treg 세포의 빈도 및 SPF 및 Hh-집락화 마우스의 MLN.
[Fig 2E] TCRHHHT 세포 및 숙주 클론의 클론형 네트워크 분석 위치(왼쪽) 및 세포 표현형(오른쪽). 완전히 연결된 각 하위 네트워크는 ‘클론형 군집’을 나타내고 각 점은 동일한 수용체 구성을 갖는 세포를 나타냅니다. 2개 미만의 세포를 갖는 클론형은 시각화를 위해 걸러냈습니다. 증식.
Myeloid compartmentalization
– 골수구획화

Fig. 3. 대식세포 개체군은 LP 미크로니체가 풍부합니다.
(A) 4개 영역에 걸친 단핵구/대식세포 및 수지상 세포 하위 집합의 매핑은 영역별로 약간의 군집링을 나타냈지만 고유한 APC 개체군은 나타내지 않았습니다
(B) 우리는 IL-1β+ 및 CD206+ 대식세포와 IL-1β+ 단핵구가 LP에서 가장 풍부했지만 LA 내에서 IL-1βhiCD103+ SIRPα+ 수지상 세포의 농축을 확인했습니다
(C-E) SIRPα+ 수지상 세포의 면역형광 염색은 LA를 조밀하게 채우는 것을 보여주는 반면(그림 3c,e), CD206 염색은 LP에 CD206+ 대식세포의 존재를 보여줍니다(그림 3d,e).
(F) 복합 MHCII 점수는 구획에 걸친 모든 골수 집단이 MHCIIhi 세포를 포함한다는 것을 시사하기 때문에 이러한 그룹과 eTreg 세포 사이에서 항원 기반 상호 작용이 발생할 수 있습니다
(G) IL-10 및 기타 STAT3 활성화 사이토카인은 장의 항상성과 염증을 제어하는 데 핵심이기 때문에 조직에서 가장 풍부한 골수 및 림프 하위 집합에 걸쳐 STAT3 의존성 사이토카인 유전자를 검색했습니다. LA에 거주하는 IL-1β+CD103+SIRPα+ 수지상 세포에서 친염증성 사이토카인 유전자 Il6의 상향 조절된 발현을 발견했습니다
[Fig. 3A] 왼쪽, 모든 위치에 걸친 총 골수성 부분집합의 UMAP 시각화. 조직 마이크로니체에 의한 총 골수성 집단의 중간, UMAP. 오른쪽, 골수성 부분집합 분포(위) 및 표시된 위치(아래)의 총 세포 수
[Fig. 3B] LA 및 LP에서 골수성, T 세포 및 ILC 비율의 log2-변환된 배수 차이
[Fig. 3C] 대표적인 CD11c 및 SIRPα 면역형광 및 Hh-감염된 맹장 조직에서의 DAPI 염색은 LA 및 주변 LP. Scale bar, 50 μm
[Fig. 3D] LA 및 주변 LP를 나타내는 Hh-감염된 맹장 조직의 대표적인 CD206 면역형광 및 DAPI 염색. Scale bars 50 μm
[Fig. 3E] Hh에 감염된 맹장 조직의 LA 및 LP 미세조직에서 CD11c+SIRPα+ 수지상 세포 및 CD206+ 대식세포의 밀도
[Fig. 3F] MHCII 발현의 바이올린 플롯은 LP(위) 및 LA(아래)에서 림프 및 골수 세포에 걸쳐 점수를 매깁니다. 영역당 30개 이상의 세포를 갖는 세포 유형으로 제한됩니다
[Fig. 3G] LA 및 LP의 골수성 및 림프성 부분집합에서 STAT3 신호전달 및 수용체와 관련된 유전자의 상대적 평균 발현. 영역당 30개 이상의 세포를 갖는 세포 유형으로 제한함
Niche disruption in colitis
대장염의 비침습성

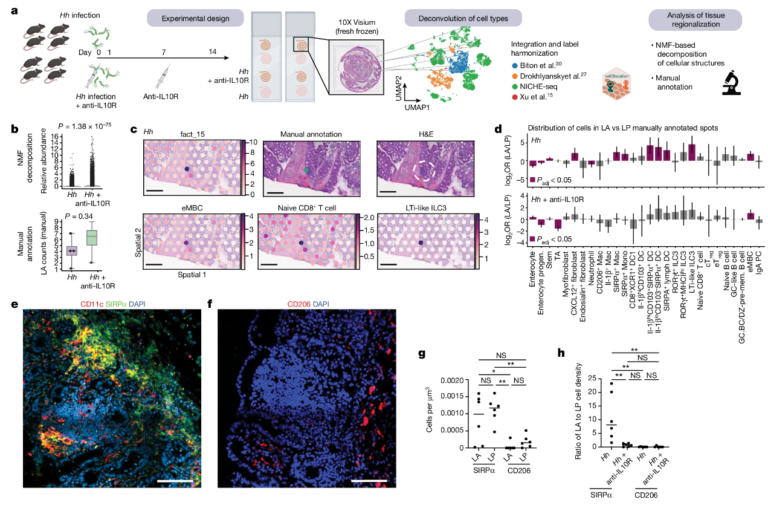
Fig. 4. 공간 전사체학 분석에 의한 LA 세포 시그니처의 농축은 염증에서 감소됩니다.
(A) 우리는 NICH-seq 데이터의 주석을 Biton et al.30, Drokhyansky et al.27 및 Xu et al.15의 데이터와 통합하고 Cell2location 파이프라인 43을 사용하여 102개의 전사적으로 구별되는 세포 유형을 조직 섹션에 공간적으로 매핑했습니다
(B) LA의 수동 주석은 NMF의 검증을 가능하게 했습니다. 두 기술 모두 대장염과 관련하여 LA 주석이 달린 조직 마이크로치가 증가하는 유사한 경향을 보여주며, 이는 아마도 LA가 분리된 림프 여포로 발달한 것을 반영한 것일 것입니다
(C) 인자 분석, 수동 주석 및 헤마톡실린 및 에오신(H&E) 염색의 예 오버레이는 LA 틈새 관련 세포 시그니처(eMBC, 나이브 CD8 및 림프 조직 유도제(LTi) 유사 ILC3)가 겹친다는 것을 보여줍니다
(D) 예상대로, 통계적으로 더 엄격한 이 분석은 IL-1βhiCD103+SIRPα+ 수지상 세포, ILC3 및 eMBC가 Hh 전용 설정의 LA에서 유의하게 풍부하다는 것을 보여주었습니다
(E-G) Hh/anti-IL10R 조건에서 SIRPα+ 수지상 세포의 면역형광 염색을 분석한 결과, 대장염의 경우 LA와 LP 사이에 유사한 분포를 보였으며(그림 4eg), 이는 세포 로딩 분석의 결과를 반영합니다. 유사하게, CD206+ 대식세포의 염색은 대장염에서 LA와 LP 사이에 유의한 차이를 보이지 않았습니다
(H) 이것은 정상 상태에서 LA에서 유의하게 풍부한 SIRPα+ 수지상 세포가 더 이상 대장염에서 유의하게 풍부하지 않다는 것을 입증했습니다
[Fig. 4A] 공간 전사체 실험의 설정 및 분석에 대한 개략도
[Fig. 4B] 상단, NMF 분석(위) 및 수동 주석(아래)에 의한 Hh/항-IL10R 마우스 대비 Hh-감염 마우스에서 LA의 상대적 존재비.
[Fig. 4C] c, Hh에 감염된 조직의 H&E 염색. 대표적인 샘플의 Visium RNA 포획 스팟에서 확장 데이터 도 8c(좌측 상단) 및 수동 주석(좌측 상단)에 정의된 바와 같은 NMF 분해에 의한 LA-연관 인자 15의 국소화. 대표적인 LA에서 eMBC(좌측 하단), 나이브 CD8+ T 세포(중간 하단) 및 LTi-유사 ILC3(우측 하단)의 국소화 및 정규화된 세포 유형 풍부. 스케일 바, 200 μm.
[Fig. 4D] 수동 주석이 달린 LA 대 Hh에 감염된 (위) 및 Hh/anti-IL10R (아래) 마우스의 LP 스팟에서 세포 계통에 따라 세포 상태 농축 순서가 지정됩니다. 통계적으로 유의한 농축(chi-square test, 조정된 P 값(Padj) < 0.05)이 마젠타에 표시됩니다. 데이터는 log2OR 값 ± s.d. 조상; TA, 전송 증폭; GC, 생식 중심; GC.BC/DZ-pre-mem, 생식 중심 B 세포/다크 존 및 사전 메모리; PC, 혈장 세포입니다.
[Fig. 4E] Hh/anti-IL10R 마우스에서 CD11c 및 SIRPα 면역형광 및 DAPI 염색, LA 및 주변 LP. Scale bar, 100 μm.
[Fig. 4F] Hh/anti-IL10R 마우스에서 CD206 면역형광 및 DAPI 염색, LA 및 주변 LP. Scale bar, 100 μm.
[Fig. 4G] Hh/anti-IL10R 마우스의 LA 및 LP에서 CD11c+SIRPα+ 수지상 세포 및 CD206+ 대식세포의 밀도.
[Fig. 4H] Hh 및 Hh/anti-IL10R 마우스의 LP 대비 LA에서 SIRPα+ 수지상 세포 및 CD206 대식세포 밀도의 비율.
eTreg cell–macrophage interactions in LP
LP에서의 eTreg 세포-대식세포 상호작용

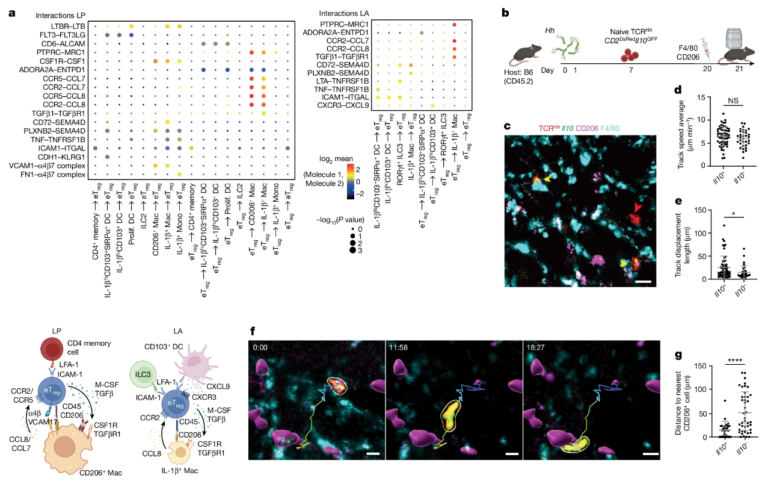
Fig. 5. in vivo live imaging은 Il10+ TCRHHT 세포가 LP에서 CD206+ = 대식세포와 상호작용하는 것을 보여줍니다.
(A) CellPhoneDB 데이터에서 강조된 CCR2-CCL8 축과 LP eTreg 세포에서 CCR2의 차등 발현(그림 5a 및 확장 데이터 그림 6i 및 9d-g)을 기반으로 eTreg 세포가 LP의 CD206+ 대식세포와 직접 상호 작용한다고 가정했습니다.
(B) 이를 테스트하기 위해 Hh-집락화된 숙주로 전달되고 F4/80 및 CD206 항체로 생체 내 표지된 TCRHhCD2DsRedIl10GFPT 세포의 생체 내 2광자 라이브 이미징을 수행했습니다
(C) LP의 이미징은 Il10- TCRHhT 세포 대응물과 대조적으로 CD206+ F4/80+ 대식세포와 밀접하게 연관된 Il10+ TCRHhT 세포를 보여주었습니다
(D-E) 유사한 세포 트랙 속도에도 불구하고 Il10-GFP+ TCRHh 세포는 더 큰 변위를 나타냈습니다
(F) 타임 랩스 이미지의 분석은 Il10-GFP+ TCRHh 세포가 한 CD206+ 대식세포에서 다른 대식세포로 이동하는 것을 보여줍니다
(G) 운동성 eTreg 세포가 LP에서 CD206+ 대식세포와 상호 작용한 결과는 Il10-GFP+ TCRHh T 세포에서 CD206+ 대식세포까지의 거리가 Il10- TCRHh 세포에 비해 상당히 작으며, 한 번에 약 1/3의 세포가 CD206+ 대식세포와 접촉한다는 것입니다
[Fig 5A] pLP(왼쪽 위) 및 LA(오른쪽 위)에서 수용체-리간드 상호작용에 대한 CellPhoneDB 분석, 영역당 30개 이상의 셀이 있는 셀 유형으로 제한되고 P < 0.01인 고유한 쌍으로 제한됩니다. MHCII 상호작용은 제외됩니다. 가장 관련성이 높은 셀-셀 상호작용을 요약한 개략도(아래).
[Fig 5B] CD206 및 F4/80 형광 항체로 생체 내에서 표지된 Hh-집락화된 숙주로 전달된 TCRHhCD2DsRedIl10GFPT 세포의 2광자 생체 내 라이브 이미징.
[Fig 5C] LP 중 공여자 TCRHhCD2DsRedIl10GFPT 세포의 대표 이미지. 화살표는 Il10- TCRHhh(빨간색) 및 Il10+ TCRHh(노란색) T세포. Scale bar, 20 μm.
[Fig 5D] LP판에 ‘일10+’와 ‘일10-TCRH-TcrH Tcells’의 비행기 속도입니다.
[Fig 5E] 트랙 배치 length of Il10+ and Il10 – TCRHT cells in the LP
[Fig 5F] LP에서 한 CD206+ 셀에서 다른 셀로 Il10+ TCRHhT 셀이 이동하는 것을 보여주는 순차 비디오 스틸, 스케일 바, 10 μm.
[Fig 5G] Il10+ 및 Il10- TCRHh에서 LP에서 가장 가까운 CD206+ 셀까지의 거리.
Disscussion
이 연구에서는 TCRHh를 미생물에 반응하는 Treg 세포의 모델로 사용하여 장에서 면역 조절 기능을 획득하고 유지하는 Treg 세포의 자연사를 추적한다. 해부학적 미세 환경에서 세포를 검토함으로써, 우리는 eTreg 세포의 항원 자극 및 효과 기능을 강화하는 핵심 상호작용의 중요성을 밝혀냈다. TCRHh T 세포를 사용하고 내인성 Treg 세포에서도 나타나는 cTreg-eTreg 분화 궤적은 미세 환경 내의 상호작용을 기반으로 한 환경적 각인을 무시하고 Treg 표현형을 주도한다는 것을 시사한다. 이는 Gata3-Rorc 이분법에 이 패러다임을 덧씌운다. eTreg 세포 중에서도 LP(라미나 프로피아)가 LA(림프 조직)가 아니라 eTreg 세포 기능의 향상된 장소이며, AREG, GZMB 및 IL-10의 생성을 포함한다.
LP는 그 고유한 세포 구성으로 안정된 상태에서 내성을 지원합니다. 이 미세 환경에는 eTreg 세포와 다양한 APC(항원 제시 세포) 집단이 서식하는데, 여기에는 CD103+ 수지상 세포, CD206+ 대식세포, IL-1β+ 대식세포, 그리고 IL10R의 가장 높은 수준을 표현하는 단핵구가 포함됩니다. 이들은 eTreg 세포가 생성하는 IL-10의 주요 대상이 됩니다. 이 내성 유도 미세 환경이 확립되면, eTreg 세포는 이 미세 환경 내에서 증식하여 병리를 조절할 수 있습니다. 그러나 염증 상황에서는 LP 미세 환경이 교란되며, 이전에 LA(림프 조직)에 국한되었던 여러 세포 유형들이 LP로 유입되는데, 이는 다시 이 조직이 점막 내성의 핵심임을 시사합니다. LP 내에서, IL-10을 생산하는 Treg 세포는 이동성이 있으며 CD206+ 대식세포와 연속적인 상호작용에 참여합니다. eTreg 세포와 CD206+ 대식세포는 이러한 상호작용을 증진할 수 있는 여러 분자를 발현하는데, 여기에는 CCR2–CCL8을 통한 유인, VCAM1–α4β7 및 CD45–CD206을 통한 접착, 그리고 TCR–MHC 및 IL-10–IL10R을 통한 면역 활성화 및 조절이 포함됩니다.
이 연구는 eTreg 세포 프로그램의 중요성과 eTreg 세포의 이동성 및 안정성을 강조하며, 이 세포들은 큰 장벽 표면을 가로질러 세포들과 상호작용할 수 있는 독특한 위치에 있습니다. 세포형태뿐만 아니라 그들의 미세 환경을 특성화함으로써, 우리는 병리를 조절할 수 있는 여러 경로들을 밝혀냈습니다. 미세 환경 연구에서 얻은 정보는 특정 개입, 세포 치료 및 백신 전략을 통해 염증성 장 조직 및 다른 장기에서 eTreg 세포의 모집, 활성화, 분화, 생존 및 기능을 지원하는 데 사용될 수 있습니다.