
IL-10은 sphingolipid metabolism 억하여 염증을 제한합니다.
Abstract
Interleukin-10(IL-10)은 innate immune cell types에서 immune cell activation과 사이토카인 생산을 제한할 수 있는 주요 anti-inflammatory cytokine입니다. IL-10 신호가 손실되면 사람과 mice에서 생명을 위협하는 inflammatory bowel disease가 발생하지만, IL-10 신호가 염증을 억제하는 정확한 메커니즘은 아직 명확하지 않습니다.
여기서 우리는 saturated very long chain (VLC) ceramides의 증가가 IL-10 deficiency의 특징인 inflammatory gene expression 증가에 중요하다는 사실을 발견했습니다. 따라서 VLC ceramides 생산을 담당하는 효소인 ceramide synthase 2 (Cers2로 암호화됨)의 genetic deletion은 in vitro and in vivo에서 IL-10 deficiency와 관련된 inflammatory gene expression이 제한되었습니다. saturated VLC ceramides의 축적은 새로운 mono-unsaturated fatty acid synthesis pathway를 통한 metabolic flux의 감소에 의해 조절되었습니다. IL-10 신호가 deficient된 세포에서 mono-unsaturated fatty acid availabIlity를 회복하면 saturated VLC ceramide production과 관련 염증이 제한됩니다. 메카니즘적으로, VLC ceramides에 의해 매개되는 persistent inflammation이 immuno-modulatory transcription factor인 REL의 지속적인 활성에 크게 의존한다는 것을 발견했습니다.
이러한 데이터를 종합하면 IL-10에 의한 fatty acid desaturation programme이 VLC ceramide 축적과 REL의 비정상적인 활성화를 일으킨다는 것을 알 수 있습니다. 이러한 연구는 innate immune cells의 fatty acid homeostasis이 병적 염증을 조절하는 key regulatory node로 작용한다는 생각을 뒷받침하며, VLC homeostasis의 ‘metabolic correction’이 IL-10의 부재로 인한 dysregulated inflammation을 정상화하는 중요한 전략이 될 수 있음을 시사합니다.
*IL-10: innate and adaptive immune cells의 immune activation을 제한하는 anti-inflammatory cytokine
Figures
IL-10은 sphingolipid metabolism을 조절.
– IL-10 신호가 TLR2 activation의 downstream에서 lipid metabolic reprogramming에 필요한지 여부를 확인
– IL-10 신호가 ceramide synthesis에 영향을 미치는지 확인
– ceramides의 축적이 TLR2-activated macrophages의 inflammatory gene expression programme에 기여하는지 여부를 테스트
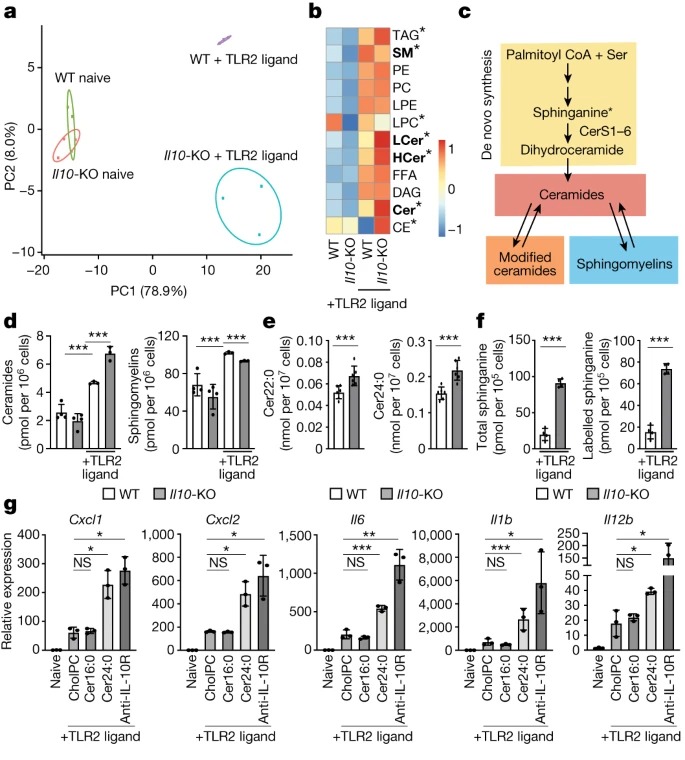
Fig. 1: IL-10 신호는 sphingolipid metabolism을 조절.
(A) 48시간 동안 naive 또는 TLR2-activated (50ng ml-1 Pam3CysK4) wild-type 및 IL10-KO BMDM에서 mass spectrometry로 정량한 individual lipids의 Principal component analysis (PCA).
(B) (A)에서와 같이 자극된 naive BMDM(왼쪽 두 열) 또는 TLR2-activated BMDMs (오른쪽 두 열)에서 direct infusion mass spectrometry로 측정한 individual lipid의 Heat map.
(C) sphingolipid metabolism의 단순화된 모식도.
(D) (A)에서와 같이 자극한 BMDM에서 direct infusion mass spectrometry로 측정한 Total ceramides and sphingomyelin species.
(E) 복강 내 주사를 통해 48 시간 TLR2 리간드 (mice 당 50μg Pam3CysK4)를 투여한 후 wild-type 및 IL10-KO mice에서 수집한 ex vivo peritoneal macrophages에서 direct infusion mass spectrometry으로 측정한 Ceramide species.
(F) 48 시간 TLR2-activated wild-type 및 IL10-KO BMDM에서 total and labelled sphinganine의 LC-MS 분석.
(G) naive BMDM과 24시간 동안 TLR2 리간드(50ng ml-1 Pam3CysK4)로 활성화된 BMDM에서 inflammatory gene expression의 Quantitative PCR(qPCR) 분석.
*fatty acid 16:0 – fatty acid chain길이 : saturated carbons 수
18:0 – stearic acid
18:1 – mono-unsaturated oleic acid
18:2 – linoleic acid
*TLR2 : toll-like receptor 2.
[Figure 1A] targeted shotgun lipidomic analysis를 사용하여 naive 또는 TLR2-activated wild-type과 IL10-knockout (KO) bone marrow-derived macrophages (BMDM)의 lipid profiles을 비교
lipidomics의 Principal component analysis 결과, naive wILd-type and IL10-KO macrophages는 principal components PC1과 PC2가 대체로 유사함. PC1은 WT 및 IL10-KO macrophages에서 TLR2-activated macrophages에서 관찰된 변이의 약 80%를 차지함. PC2(변이의 8%)는 TLR2 downstream의 lipid composition에 대한 IL-10 신호의 영향을 묘사.
[Figure 1B-D] TLR2 활성화 후 IL-10 신호의 영향을 받는 lipids를 자세히 살펴보면 ceramides, modified ceramides (hexosyl ceramides and lactosyl ceramides), sphingomyelins의 축적이 두드러지게 나타났고 sphingomyelins은 TLR2 활성화 후 감소.
[Figure 1E] in vitro 결과와 일치하게, IL-10 deficient ex vivo macrophages는 많은 ceramide species이 유의하게 증가하고 mono-unsaturated sphingomyelin species이 유의하게 감소하는 것으로 나타남.
– 따라서 IL-10 신호가 in vitro and in vivo inflammatory stimuli에 대한 반응으로 macrophages의 sphingolipid metabolism을 조절한다는 결론내림.
[Figure 1F] IL-10 신호가 deficient된 macrophages의 Targeted liquid chromatography mass spectrometry (LC-MS)은 wild-type controls에 비해 total 및 동위원소 표지된 sphinganine을 더 많이 함유함.
– 이는 IL-10 신호가 부족한 macrophages에서 관찰된 ceramides의 이상 축적에 de novo ceramide synthesis 증가가 기여함을 나타냄.
[Figure 1G] cholesteryl-phosphocholine으로 구성된 lipid bilayers를 만들어, 비어 있거나(CholPC로 표시) ceramide 16:0(가장 풍부한 long chain ceramide인 Cer16:0) 또는 ceramide 24:0(가장 풍부한 VLC ceramide인 Cer24:0)과 complexed 시킴.
TLR2-activated wild-type macrophages에 Cer16:0을 추가해도 inflammatory gene expression에 변화가 없었음. 반면, Cer24:0을 추가하면 macrophages viability에 영향을 주지 않으면서 inflammatory gene expression이 IL-10/IL-10R 신호가 없을 때와 유사한 수준으로 상향 조절됨.
– 이러한 데이터를 종합하면 VLC ceramides의 축적이 TLR-activated macrophages에서 cytokine and chemokine gene expression을 향상시킬 수 있으며, dysregulation of VLC ceramide homeostasis가 IL-10 deficiency에서 관찰되는 염증 악화에 기여한다는 것을 시사.
VLC ceramides의 loss는 inflammation을 제한.
– CerS2와 그 sphingolipid products가 inflammation에 중요한지 확인.
– CerS2가 IL10-KO 또는 IL10rb-KO mice의 특징인 colonic inflammation을 조절하는 데 중요하다는 가설 확인.
– CerS2가 colonic homeostasis에 미치는 영향 확인.
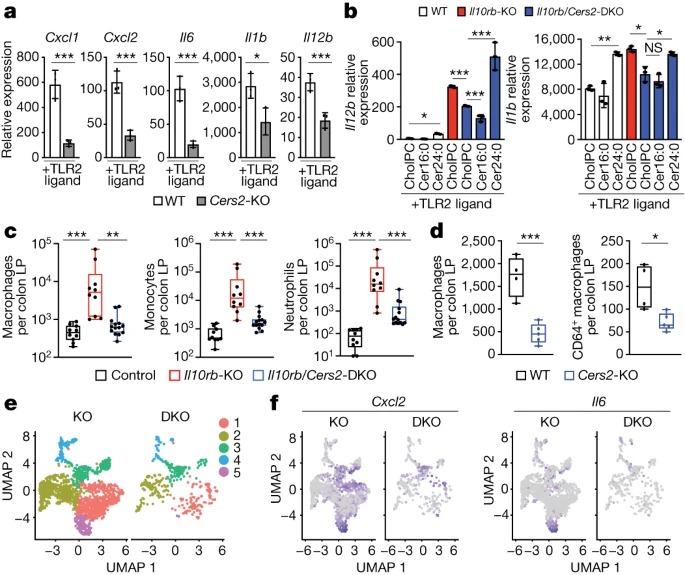
Fig. 2: VLC ceramide synthesis의 Genetic inhibition은 inflammation을 제한함.
(A) 48시간 TLR2 활성화(50ng ml-1 Pam3CysK4) wild-type 또는 Cers2-KO BMDM에서 Inflammatory gene expression의 qPCR 분석.
(B) 활성화 마지막 44시간 동안 cholPC(vehicle), Cer16:0 또는 Cer24:0을 보충한 48시간 TLR2-activated (50ng ml-1 Pam3CysK4) wild-type, IL10rb-KO 또는 IL10rb/Cers2-DKO peritoneal macrophages에서 Il12b 및 Il1b 유전자 발현의 qPCR 분석.
(C) 대조군(IL10rb-heterozygous), IL10rb-KO 및 IL10rb/Cers2-DKO 키메라 mice의 colonic lamina propria (LP)의 flow cytometry-based immune cell profiling에서 macrophages(CD11c+MHCII+), monocytes (CD11b+Ly6C+) 및 neutrophils (Cd11b+Ly6G+) 측정.
(D) wild-type 및 Cers2-KO 키메라 mice의 colonic lamina propria에서 나온 Total macrophages (CD11c+MHCII+) 및 CD64+ macrophages.
(E) colonic lamina propria에서 분류된 세포의 IL10rb-KO 및 IL10rb/Cers2-DKO macrophages 클러스터에서 single-cell RNA-seq(scRNA-seq) 데이터의 UMAP 분석.
(F) IL10rb-KO 또는 IL10rb/Cers2-DKO 키메라 mice colonic lamina propria에서 분류된 macrophages의 Cxcl2 및 Il6 scRNA-seq 데이터에 대한 UMAP 분석.
[Figure 2A] Cers2-heterozygous macrophages는 wild-type counterparts와 유사한 inflammatory gene expression을 보임. 반면, Cers2-KO macrophages는 TLR2 ligands에 반응하여 inflammatory gene expression이 현저히 감소했으며, IL-10R 차단 시 inflammatory gene expression이 완전히 유도되지 않았음.
– 이는 CerS2 기능이 TLR2-driven macrophage inflammation 단독 또는 IL-10 deficiency의 맥락에서 중요하다는 것을 시사.
[Figure 2B] IL-10R deficiency 배경에서 CerS2를 제거하면 TLR2 및 TLR4 리간드에 반응하여 macrophage inflammatory gene expression이 현저히 감소. 특히, Cer16:0이 아닌 exogenous Cer24:0을 추가하면 IL10rb/Cers2-DKO macrophage의 inflammation이 회복됨.
– 이는 CerS2의 lipid products인 VLC ceramides의 축적이 IL-10-deficient macrophages의 inflammatory gene expression을 매개하는 데 중요하다는 것을 나타냄.
[Figure 2C] CD45.2+ IL10rb+/-(control) mice, IL10rb-KO mice, IL10rb/Cers2-DKO littermate mice에서 CD45.1+ recipients로 bone marrow 키메라를 만듬.
IL10rb-KO 키메라에서는 macrophages, monocytes, neutrophils, total CD4+ T cells, TH1 (IFNγ+) and TH17 (IL-17A+) cells을 포함한 inflammatory cell infiltrates이 증가.
IL-10R-deficient background에서 CerS2가 deficient된 경우 IL10rb-KO 키메라에 비해 inflammatory cell infiltrates이 현저히 감소함.
– 이는 IL-10 신호가 없을 때, immune cell-mediated colitis를 유발하는 데에 VLC ceramides의 합성이 중요하다는 것을 나타냄.
[Figure 2D] Cers2-KO 키메라 mice는 macrophages (CD11c+MHCII+)와 mature macrophages (CD11c+MHCII+CD64+)가 감소한 것으로 나타남.
– basal conditions에서 CerS2가 colonic macrophage populations의 유지에 중요하다는 것을 시사.
[Figure 2E,F] Uniform manifold approximation and projection (UMAP) 분석 결과, IL10rb-KO 키메라에서 분리한 colonic macrophages는 5개의 별개의 클러스터로 그룹화되었으며 클러스터 5에는 inflammatory cells이 많이 포함됨.
특히, IL10rb/Cers2-DKO 키메라에는 클러스터 5가 없었으며, IL10rb-KO 키메라에 비해 inflammatory cells markers, cytokines and chemokines의 발현이 감소함.
또한 CerS2-deficient macrophages는 CD206(encoded by Mrc1) 및 기타 tolerogenic markers의 높은 발현을 보였음.
– 이는 CerS2의 loss가 IL-10R 단독 loss에 비해 더 tolerogenic macrophage phenotype을 촉진할 수 있음을 시사.
– 이러한 데이터를 종합하면, IL-10R deficient background에서 CerS2의 loss는 IL-10R 단독 loss에 비해 immune cell의 colon infiltrates를 완화하고 in vivo macrophage inflammatory gene expression을 감소시킬 수 있음을 시사.
MUFA 합성은 ceramides 생산을 제한.
– IL10-KO 세포에서 ceramides 합성이 어떻게 조절되는 확인
– IL10-KO macrophages에서 관찰된 Scd2 gene expression 감소가 de novo MUFA 합성을 변화시키기에 충분한지 확인.
– IL10-KO BMDM의 변화된 MUFA 함량이 염증에 중요한지 확인
– IL10-KO BMDM에서 mono-unsaturated oleic acid(18:1)의 첨가가 ceramide homeostasis에 어떤 영향을 미치는지 조사
– in vivo inflammation에 대한 SCD2의 영향을 확인.
– Scd2-cKO mice에서 발견되는 basal inflammation 증가가 이 mice가 dextran sodium sulfate (DSS)과 같은 chemically-induced colitis 모델에 취약할 수 있다는 가설.
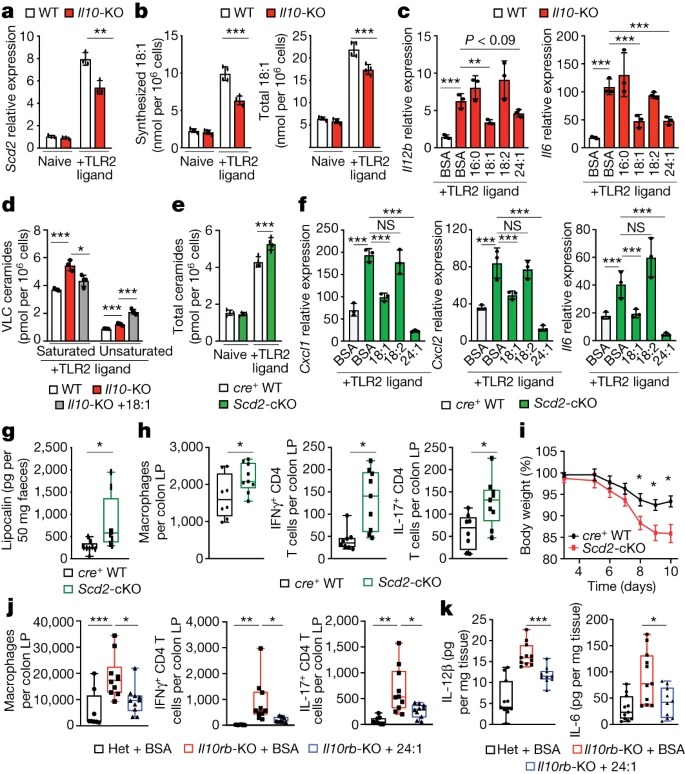
Fig. 3: IL-10에 의한 mono-unsaturated fatty acid synthesis는 ceramide production을 제한.
(A) naive 또는 24 시간 TLR2-activated wild-type 또는 IL10-KO BMDM에서 Scd2 gene expression의 qPCR 분석.
(B) naive 또는 48 시간 TLR2-activated wild-type 또는 IL10-KO BMDM에서 Synthesized and total oleic acid.
(C) 지난 44시간 동안 BSA 또는 25μM BSA-16:0, BSA-18:1, BSA-18:2 또는 BSA-(nervonic acid)24:1로 배양한 48시간 TLR2-활성화 wild-type 또는 IL10-KO BMDM에서 Il12b 및 Il6 유전자 발현의 qPCR 분석.
(D) 마지막 44시간 동안 25μM 18:1로 처리한 48시간 TLR2-activated wild-type 또는 IL10-KO 또는 IL10-KO BMDM의 Saturated 및 unsaturated ceramides 종.
(E) naive 또는 TLR2-activated LysM-cre+ wild-type 및 Scd2-cKO BMDM의 Total ceramides species.
(F) BSA로 배양한 48시간 TLR2-activated cre+ wild-type BMDM, BSA로 배양한 Scd2-cKO BMDM 및 마지막 44시간 동안 25μM BSA-18:1, BSA-18:2 또는 BSA-24:1로 배양한 Scd2-cKO BMDM에서 Cxcl1, Cxcl2 및 Il6 유전자 발현의 qPCR 분석.
(G) 대조군 및 Scd2-cKO 수컷 mice의 faecal lipocalin에 대한 Enzyme-linked immunosorbent assay (ELISA) 분석.
(H) naive 대조군 및 Scd2-cKO 수컷 mice의 colonic lamina propria의 Flow cytometry-based immune cell profiling.
(I) 대조군 및 Scd2-cKO 암컷 mice에서 DSS-induced weight loss.
(J) BSA를 투여한 IL10rb- heterozygous (Het) 수컷 mice와 BSA 또는 BSA-24:1을 투여한 IL10rb-KO 수컷 mice의 colonic lamina propria의 Flow cytometry-based immune cell profiling.
(K) ex vivo에서 24시간 동안 배양한 BSA 또는 BSA-24:1로 배양한 IL10rb- heterozygous mice 및 IL10rb-KO 수컷 mice의 colonic explants에서 IL-6 및 IL-12β의 ELISA 분석.
*stearoyl-CoA desaturase 2 (Scd2) : fatty acid의 double bond 형성을 촉진시키는 효소.
*MUFAs : mono-unsaturated fatty acids
*faecal lipocalin : 대장에서 염증과 비례하는 병리학적 지표.
[Figure 3A] genotype이나 activation status에 따른 sphingolipid gene mRNA 수준에는 차이가 없었지만, IL10-KO BMDM에서 stearoyl-CoA desaturase 2 (Scd2)의 발현이 유의하게 감소하는 것을 관찰.
[Figure 3B] TLR2 자극이 wild-type BMDM에서 de novo lipogenesis를 증가시켜 18:1의 합성과 총량이 증가한다는 것을 발견함. IL-10이 deficient된 Macrophages는 TLR2의 downstream에서 MUFA 합성을 완전히 상향 조절하지 못하여 자극 48시간 후 18:1의 합성과 총량이 감소함. 반면 palmitate (16:0) 합성은 변하지 않았고, imported polyunsaturated fatty acid인 linoleic acid(18:2)와 16:0의 total cellular amounts은 감소하지 않았음.
[Figure 3C] 18:1를 첨가하여 total 18:1를 약 2배 증가시킨 결과, IL10-KO 또는 IL10rb-KO BMDM에서 inflammatory gene expression이 감소함. 또한 inflammatory gene expression의 elongation product인 24:1를 첨가하면 IL10-KO BMDM에서 inflammatory gene expression이 감소한 반면 16:0및 18:2는 그렇지 않았음.
[Figure 3D] 18:1을 첨가해도 IL10-KO macrophage의 total ceramides 농도는 낮아지지 않았지만, Cer24:0을 포함한 saturated VLC ceramides의 수준은 크게 낮아지는 것을 확인. 특히 18:1를 첨가하면 Cer24:1과 total mono-unsaturated ceramides의 양이 증가함.
– 이는 saturated VLC ceramides만이 inflammatory phenotype을 유발할 수 있음을 나타냄.
[Figure 3E] IL10-KO BMDM과 유사하게, Scd2-cKO BMDM은 synthesized Cer16:0, total Cer16:0 및 total 18:2 lipid species에 영향을 미치지 않고 synthesized 및 total 18:1가 유사하게 감소. 특히, Scd2-cKO macrophages는 IL10-KO와 유사하게 sphinganine의 synthesis 증가와 total 및 saturated VLC ceramides의 축적을 보였음.
[Figure 3F] 18:1 또는 24:1를 추가하면 18:2가 아닌 TLR2 활성화에 반응하여 Scd2-cKO macrophages에서 inflammatory gene expression이 증가하는 것을 관찰함.
– 이러한 연구 결과를 종합하면, MUFA 합성을 상향 조절하지 못하는 것이 IL-10 deficiency에서 saturated ceramides의 축적과 inflammation 증가의 주요 근본 원인이라는 것을 시사.
[Figure 3G] in vitro data와 일치하게, Scd2-cKO mice는 cohoused cre+ littermate controls에 비해 faecal lipocalin의 basal elevation으로 나타남.
[Figure 3H] Scd2-cKO mice의 colon에서 macrophages의 수가 증가했으며 neutrophil 또는 monocyte 개체군에는 차이가 없었음. Scd2-cKO mice는 또한 IFNγ+ 및 IL-17A+CD4+ T 세포의 수가 증가함.
– 이는 myeloid-specific SCD2 expression이 colon에서의 T cell activation에 영향을 미친다는 것을 나타냄.
[Figure 3I] DSS-treated Scd2-cKO mice는 cohoused littermate cre+ control mice보다 훨씬 더 많은 체중이 감소하고 결장 내층에 침윤하는 myeloid 및 림프 lymphoid immune 세포의 수가 유의하게 증가.
– SCD2-deficient macrophages와 mice에서 관찰된 표현형은 IL-10 신호가 deficient된 macrophages와 mice의 표현형과 같았음.
[Figure 3J,K] MUFA를 투여한 mice는 BSA만 투여한 mice에 비해 faecal lipocalin이 현저히 감소. 또한, MUFA 섭취는 결장에서 myeloid 및 lymphoid immune cell 집단을 감소시키고 colonic explants에서 IL-6 및 IL-12p40의 분비를 감소시킴.
– 이는 CerS2 제거 결과와 유사한 수준.
반대로, Cer16:0 투여는 wild-type 또는 IL10rb-KO mice의 myeloid cell infiltration에 영향을 미치지 않음.
– 이러한 데이터는 macrophages에서 MUFA 합성의 상향 조절이 염증을 완화하는 IL-10 신호의 downstream에서 negative feedback mechanism으로 작용하며, macrophages에서 MUFA availability가 intestinal inflammation을 조절하는 데 중요한 역할을 한다는 생각을 뒷받침함.
REL은 ceramide-induced inflammation에 필요.
– VLC ceramides의 증가는 IL-10 deficiency의 특징인 late-stage inflammatory에만 영향을 미칠 수 있다는 가설.
– saturated VLC ceramides(예: Cer24:0)가 REL 활동을 구체적으로 조절하여 inflammation에 영향을 미친다는 가설
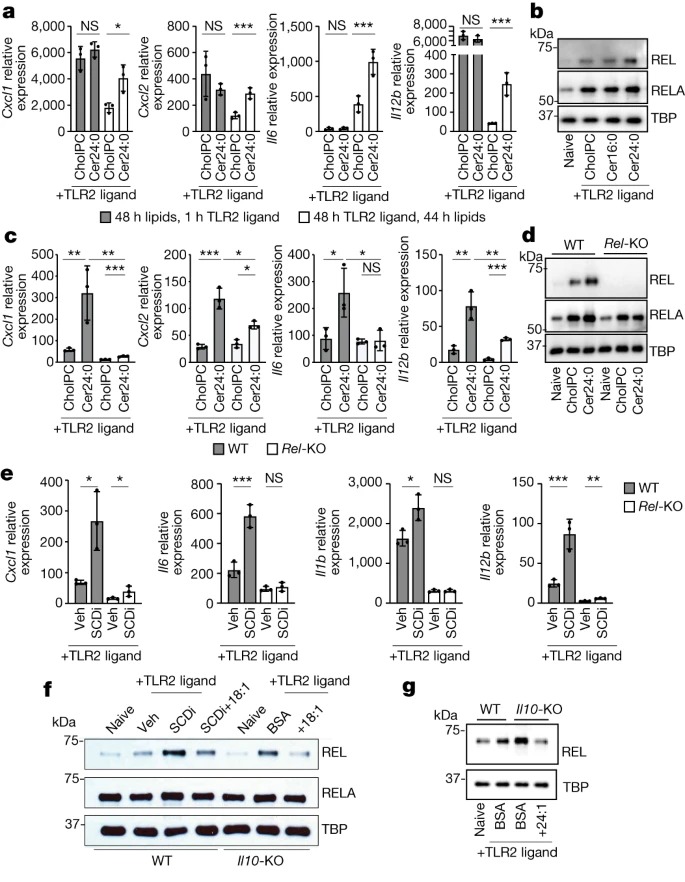
Fig. 4: REL은 inflammatory gene expression의 ceramide-mediated induction에 필요.
(A) 48시간 동안 30 μM Cer24:0으로 전처리한 후 TLR2 리간드(50 ng ml-1 Pam3CSK4)로 1시간 활성화한 BMDM에서 inflammatory gene expression의 qPCR 분석과 44시간 30 μM Cer24:0 처리로 48시간 TLR2 활성화 비교.
(B) 활성화 마지막 44시간 동안 naive 또는 48시간 TLR2-activated wild-type peritoneal macrophages의 nuclear extracts과 cholPC(vehicle), Cer16:0 또는 Cer24:0을 추가한 후의 RELA, REL 및 TBP(loading control)의 Western blot analysis.
(C) 활성화 마지막 44시간 동안 48시간 TLR2-activated wild-type 또는 Rel-KO BMDM에 cholPC(vehicle) 또는 30μM Cer24:0을 추가한 후 inflammatory gene expression의 qPCR 분석.
(D) 활성화 마지막 44시간 동안 naive 또는 48시간 TLR2-activated wild-type 또는 Rel-KO BMDM에 cholPC(vehicle) 또는 30μM Cer24:0을 추가한 후 nuclear extracts에서 RELA, REL 및 TBP(loading control) Western blot analysis.
(E) 활성화 마지막 44시간 동안 10nm SCDi(Cay10566) 유무에 관계없이 48시간 TLR2-activated wild-type 또는 Rel-KO BMDM에서 inflammatory gene expression의 qPCR 분석.
(G) 활성화 마지막 44시간 동안 naive 또는 48시간 TLR2-activated wild-type 또는 IL10-KO BMDM과 SCDi, BSA 또는 25μM BSA-18:1의 nuclear extracts에서 REL, RELA 및 TBP(loading control)의 Western blot analysis.
(F) 활성화 마지막 44시간 동안 naive 또는 48시간 TLR2-activated wild-type 또는 IL10rb-KO BMDM에 BSA 또는 25μM BSA-24:1을 추가한 후 nuclear extracts에서 RELA, REL 및 TBP(loading control) Western blot analysis.
*REL, RELA : NF-κB (Nuclear Factor-kappa B) family
[Figure 4A] Cer24:0으로 전처리해도 inflammatory genes의 초기 유도에는 변화가 없었지만 이후 시점에서는 발현이 더 오래 지속됨.
[Figure 4B] Cer16:0 처리가 아닌 Cer24:0 처리는 24시간 및 48시간 TLR2 활성화 후에 IκBα, REL 또는 RELA의 total cellular amount에 영향을 미치지 않고 nuclear REL 및 RELA가 증가했으며 이는 IL-10 deficiency의 결과와 유사
[Figure 4C] Rel-KO BMDM은 TLR2 또는 TLR4 리간드로 활성화되었을 때 VLC ceramides에 반응하여 inflammatory gene expression이 감소하는 것으로 나타남.
endogenous ceramide levels과 exogenous ceramide uptake는 REL deficiency에 의해 변화하지 않았으며, 이는 REL이 ceramide-induced inflammation의 downstream에 있음을 시사.
[Figure 4D] REL의 손실은 Cer24:0을 매개로 한 RELA의 유도를 막는 것을 확인.
– 이는 VLC ceramides의 pool size가 REL을 직접 조절할 수 있으며, enhanced RELA nuclear localization은 REL과의 dimerization에 의존하거나 REL 활성화의 downstream신호에 의해 매개된다는 것을 암시하는 결과임.
[Figure 4E,F] inflammatory genes의 SCDi-mediated induction이 REL에 의존하며, SCDi 처리 또는 Scd2의 조건부 녹아웃이 macrophages에서 REL nuclear localization을 유도할 수 있으며, 이는 exogenous 18:1를 추가함으로써 완화될 수 있음을 발견.
[Figure 4F,G] IL10-KO 또는 IL10rb-KO macrophages에서 REL의 nuclear localization은 18:1 (Fig. 4f) 또는 24:1 (Fig. 4g)의 첨가에 의해 제한되는 반면, RELA의 localization 변화는 영향을 덜 받았음.
– 이러한 데이터를 종합하면 REL은 VLC ceramide-mediated induction of inflammatory genes에 필수적인 역할을 하며 IL-10 또는 MUFA-deficient mice와 macrophages에서 발견되는 enhanced inflammatory phenotypes에 주로 관여한다는 것을 확인.
Disscussion
이 연구에서 우리는 immune response, lipid metabolism and intestinal homeostasis의 알려진 transcriptional regulators 간의 crosstalk을 메카니즘적으로 설명합니다. 여기서 우리는 anti-inflammatory cytokine IL-10이 염증의 지속 시간을 조절하는 이전에 설명되지 않은 메커니즘을 밝혀냈습니다. 특히 IL-10 신호가 없는 경우, de novo sphingolipid synthesis pathway로의 유입이 증가하여 inflammatory gene expression을 촉진하는 VLC ceramides가 축적되는 것을 발견했습니다. 예상치 못하게도 sphingolipid production의 변화는 합성 경로에서 enzymes을 암호화하는 유전자의 transcript levels에 의해 매개되지 않는다는 사실을 발견했습니다. 대신, ceramides biogenesis의 변화는 IL-10이 Scd2 유전자 발현과 그에 따른 long chain MUFAs의 합성을 제어하는 능력으로 추적되었습니다. 특히, SCD2의 pharmacologic inhibition or genetic deletion은 IL-10 결실과 유사한 방식으로 새로운 ceramides 합성 경로에 관여했습니다. de novo ceramides 합성 경로로의 유입을 조절하는 알려진 체크포인트는 거의 없으며, de novo MUFA 합성이 이 경로를 어떻게 길항하는지는 아직 명확하지 않습니다. 한 가지 가능성은 MUFA 합성이 제한되면 palmitate와 같은 saturated fatty acid precursors가 증가한다는 것입니다. palmitoyl-CoA와 serine의 condensation은 ceramides de novo 합성 경로의 첫 번째 단계이며 모든 downstream sphingolipid metabolites의 생성에 필요합니다. 실제로 IL-10이 deficient되면 macrophages에서 total cellular palmitate가 약간 증가합니다. 그러나 MUFA synthesis를 둔화시키고 total ceramides를 증가시키는 SCD2의 genetic deletion은 total 또는 synthesis palmitate에 영향을 미치지 않아, 다른 조절 메커니즘이 더 가능성이 높다는 것을 시사합니다. 특히, SCDi 처리 또는 Scd2의 conditional knockout은 N-acylated tail의 saturation에 관계없이 대부분의 ceramides species의 양을 증가시키는 것으로 나타났습니다. 이는 MUFA가 ceramides acyl tail과 관련하여 제한적이지 않으며 substrate availability 외부에서 ceramides biogenesis로의 플럭스를 조절하는 데 더 광범위한 역할을 할 수 있음을 시사합니다. 따라서 또 다른 가능성은 IL-10-mediated MUFA synthesis가 orosomucoid (ORM) 계열 단백질의 활성에 관여할 수 있다는 것입니다. ORM 단백질은 SPTLC2와 같은 serine palmitoyl transferases와 길항하여 sphingolipid 경로로의 플럭스를 감소시킵니다. 특히 ORM은 airway inflammation에 영향을 미치는 것으로 알려져 있으며 다른 대사 프로그램에 의해 조절되는 것으로 보고되었습니다. IL-10 신호와 MUFAs가 ceramide de novo synthesis에 미치는 영향에 필요한 인자를 완전히 조사하기 위해서는 추가적인 생화학적 분석이 필요할 것입니다.
ceramides가 염증과 세포 사멸에 미치는 영향을 조사한 많은 이전 연구에서는 포유류 세포에서 endogenously하게 합성되지 않는 short chain C2 ceramides 유사체를 사용했습니다. 여기서는 면역 세포에서 endogenously하게 생성되는 long chain 및 VLC ceramides에 초점을 맞추고, C2 ceramides와 달리 long chain 및 VLC ceramides의 exogenous addition은 세포 사멸을 유도하거나 inflammasome 활성화를 촉진하지 않는 것으로 나타났습니다. 또한 long chain ceramides가 아닌 saturated VLC ceramides만이 macrophages 염증을 강화할 수 있음을 관찰하여(Fig. 1g), silico에서 예측한 결과에 대한 실험적 증거를 제시했습니다. 또한, VLC ceramides 꼬리의 ‘desaturation status’가 macrophages 염증의 크기를 조절하는 데 중요한 추가 기능임을 확인했습니다. 이러한 연구 결과는 VLC ceramides가 inflammation machinery에 정보를 전달하는 방식에 대한 놀라운 수준의 특이성을 시사하며, 향후 연구에서는 unsaturated ceramides가 아닌 saturated ceramides가 inflammatory gene expression을 매개하는 방식에 초점을 맞추는 것이 중요할 것입니다. 또한, downstream sphingolipid metabolites가 염증에 어떤 영향을 미치는지 알아보기 위한 추가 연구가 필요합니다. 따라서 macrophages에서 IL-10 신호와 sphingolipidome의 역할을 완전히 규명하기 위해서는 추가 연구가 필요합니다. 또한 IL-10 신호의 손실은 대부분의 saturated 및 모든 mono-unsaturated sphingomyelins을 감소시킨다는 점에 주목할 필요가 있습니다(Fig. 1d). 이는 MUFA의 감소(Fig. 3b), sphingomyelin synthesis에 필요한 endoplasmic reticulum-to-Golgi transport 장애 또는 sphingomyelin degradation증가로 인한 것일 수 있습니다. immune cell types에서 inflammatory cytokine signalling에 대한 반응으로 강화된 sphingomyelin degradation이 관찰되었지만, sphingomyelin degradation 증가가 염증에 어떤 영향을 미치는지에 대한 정확한 메커니즘은 아직 잘 알려지지 않았습니다. IL-10 신호가 sphingolipid salvage pathways에 영향을 미치는지 여부와 그 방법을 규명하기 위해서는 추가 연구가 필요합니다.
우리는 VLC ceramides에 대한 response 또는 MUFA synthesis의 부재에서 관찰되는 persistent inflammation이 NF-κB 가족 구성원인 REL의 지속적인 활동으로 추적될 수 있음을 발견했습니다. IL-10과 REL 간의 Crosstalk은 이전에 macrophages에서 연구된 바 있지만, REL transcriptional activity가 IL-10 신호의 downstream fatty acid desaturation에 의해 영향을 받을 수 있다는 관찰은 fatty acid homeostasis manipulation이 immune cell activation과 직접적으로 연관되어 있다는 생각을 강화합니다. 또한 REL이 IL-10 신호가 없을 때 지속적인 inflammatory gene expression에 중추적이며 myeloid cells의 원형 표적인 Il12a 및 Il12b 외부의 inflammatory gene expression에 중요하다는 강력한 증거를 제공합니다. 따라서 REL 활성을 조절하는 lipid sensor signalling pathways를 규명하는 것은 광범위한 metabolic 및 sterile inflammatory diseases에서 관찰되는 비정상적인 염증을 조절하는 새로운 표적으로 작용할 수 있습니다. saturated VLC ceramides가 REL cellular location을 구체적으로 지시하는 정확한 메커니즘은 아직 명확하지 않으며 향후 연구의 중요한 주제가 될 것입니다. VLC ceramides에 대한 반응으로 REL nuclear location이 RELA보다 더 크게 영향을 받는 이유도 아직 명확하지 않은데, 이는 inflammatory kinetics, transcription factor-specific post-translational modifications 또는 특정 NF-κB 계열을 표적으로 하는 알려지지 않은 신호 캐스케이드 때문일 수 있습니다. VLC ceramides가 REL 활성화에 어떻게 관여하는지 완전히 이해하려면 추가 연구가 필요합니다.
마지막으로, VLC ceramides의 유전적 감소 또는 MUFA를 경구 투여하면 IL-10 또는 IL-10R-deficient mice에서 발견되는 colonic inflammation을 줄일 수 있음을 발견했습니다(Fig. 2c-f). 이는 de novo lipid synthesis pathways의 조절이 colonic homeostasis 및 건강 유지에 중요함을 시사합니다. IL-10 또는 IL-10R에 deleterious mutations이 있는 사람은 생후 몇 개월 이내에 생명을 위협하는 심각한 장염이 발생합니다. 이러한 사람들은 anti-TNF biologicals이나 스테로이드와 같은 표준 형태의 면역 억제에 효과적으로 반응하지 않습니다. haematopoietic stem cell transplant 외에는 기능적 IL-10 신호가 부족한 환자의 IBD에 효과적인 치료 옵션이 없습니다. 흥미롭게도 unsaturated fats이 많은 지중해식 식단을 통한 식이 요법은 6개월 만에 IBD 환자의 질병 매개 변수를 감소시키는 것으로 나타났습니다. 지중해식 식단의 핵심 구성 요소는 MUFA가 풍부한 올리브 오일입니다. 따라서 지중해식 식단의 이점은 부분적으로는 VLC ceramides 대사의 변화를 통한 MUFA-mediated repression of colonic inflammation에 의존할 수 있다고 추측할 수 있습니다. 실제로 당사의 mice 데이터는 exogenous MUFA의 경구 전달이 colon의 inflammation을 유익하게 억제할 수 있으며 fatty acid ‘metabolite correction’에 초점을 맞춘 새로운 대장염 치료법을 확립하는 데 도움이 될 수 있음을 강력하게 시사합니다.