
Annelid functional genomics는 bilaterian life cycle의 기원을 밝힌다.
Abstract
intermediate larva의 간접적인 발달은 모든 주요 동물 계통에 존재하며, 이는 larvae를 동물 진화의 대부분의 시나리오의 중심으로 만든다. 하지만 larvae가 어떻게 진화했는지는 여전히 논란의 여지가 있다.
여기서 우리는 몸통 formation의 시간적 변화(즉, heterochronies)가 larvae와 bilaterian의 생활 주기의 다양화를 뒷받침한다는 것을 보여준다. 우리는 이것과 다른 두 개의 환형체의 수명 주기 동안 transcriptomic and epigenomic profiling을 사용하여 환형체 Owenia fusiformis에서 chromosome-scale genome sequencing을 수행했다. 우리는 몸통 development가 O.fusiformis의 feeding larva에서 pre-metamorphic stage로 지연되지만, Capitella teleta의 점진적인 metamorphosis와 Dimorphilus gyrociliatus의 직접적인 발달 배아와 함께 non-feeding larva의 gastrulation 이후에 시작된다는 것을 발견했다. 따라서 O. fusiformis의 배아는 먼저 larval 조직과 성체의 머리를 형성하는 enlarged anterior domain으로 발달한다. 특히, 이것은 다른 bilaterian의 이른바 ‘head larvae’에서도 발생하는데, O. fusiformis larva와 광범위한 transcriptomic 유사성을 보인다.
우리의 연구 결과는 head larvae에서 최대로 관찰된 것처럼 머리와 몸통 형성의 시간적 분리가 Bilateria에서 larval evolution을 촉진했음을 시사한다. 이것은 larvae와 성인 기원을 설명하기 위해 유전자 조절 프로그램의 co-option 또는 innovation을 제안하는 일반적인 시나리오와는 다르다.
Figure
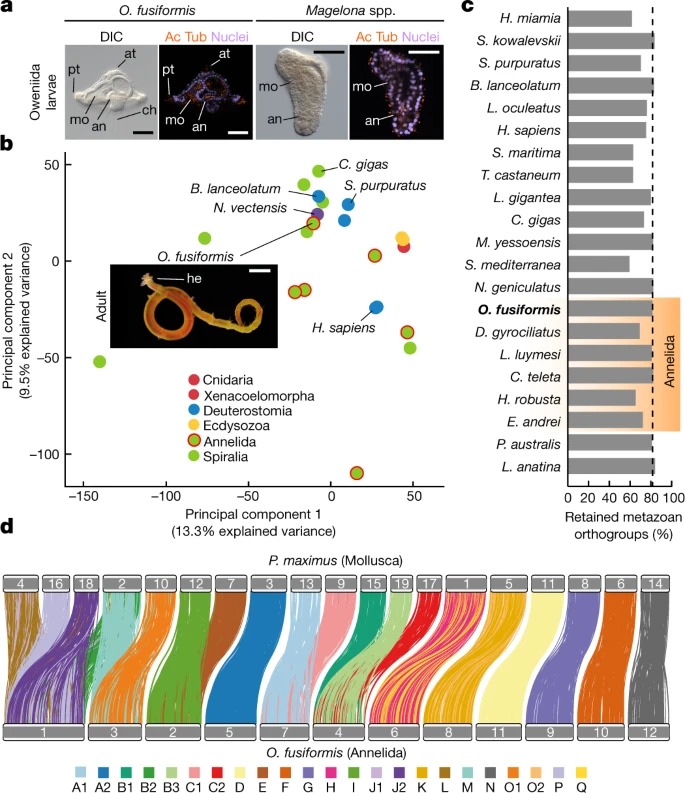
Fig. 1. O. fusiformis는 구분되는 larva와 conservatively 진화하는 게놈을 가지고 있다.
a, oweniids and magelonid의 larvae는 다른 환형동물의 larvae와는 다르다. O. fusiformis mitraria 및 Magelona spp의 DIC (Differential interface contrast) 이미지 및 z-stack confocal 레이저 스캔.
b, metazoan gene complement의 Principal component analysis 은 conservatively 진화하는 유전자를 가진 다른 계통의 O. fusiformis 클러스터가 보완한다는 것을 보여준다.
c, 종당 pre-metazoan and metazoan orthogroups의 보존 비율.
d, O. fusiformis과 Pecten maximus 사이의 Karyotypic correspondence는 조상의 spiralian chromosome complement의 예시이다.
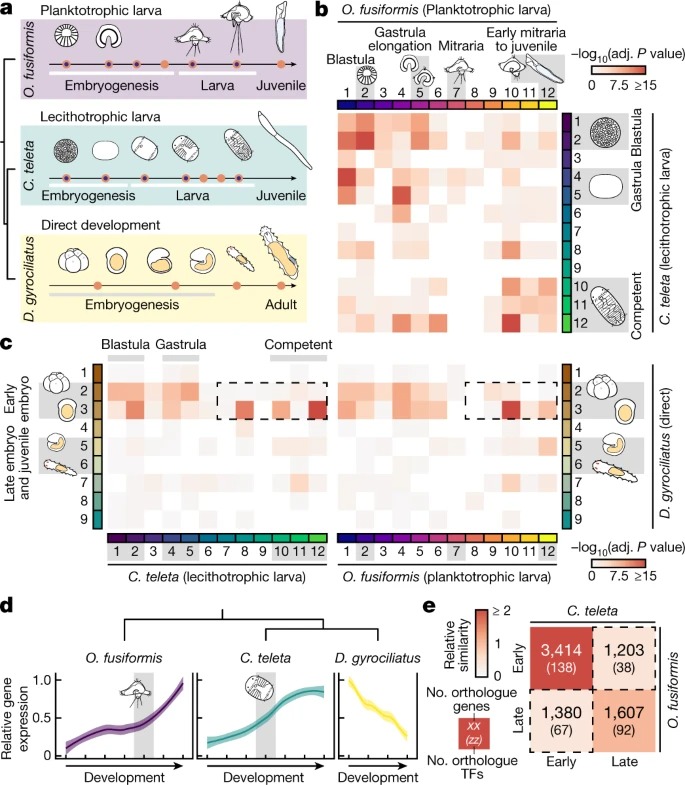
Fig. 2. gene regulatory programmes의 heterochrony은 annelid life cycle 다양화를 뒷받침한다.
a, comparative developmental RNA sequencing (RNA-seq)의 실험 설계 및 assay for transposase-accessible chromatin with sequencing (ATAC-seq) time courses.
b,c, 연구된 세 개의 환형 분류군(O. fusiformis과 C. teleta의 경우 12개 클러스터, D. gyrociliatus의 경우 9개 클러스터) 사이의 soft k-means clustering에 의해 얻은 co-regulated gene 클러스터에 포함된 transcription factor 사이의 orthogroup overlap을 보여주는 Similarity heatmap.
d, 28개의 single-copy orthologue transcription factor의 Average expression dynamic는 O. fusiformis과 C. teleta 모두에서 late expression에서 D. gyrociliatus의 early expression으로 전환되었다.
e, O. fusiformis and C. teleta에서 pre-larval (early) 클러스터와 post-larval (late) 클러스터 사이의 사분면에 의한 whole-genome orthogroup overlap analysis에 기초한 상대적 유사성의 Heatmap.
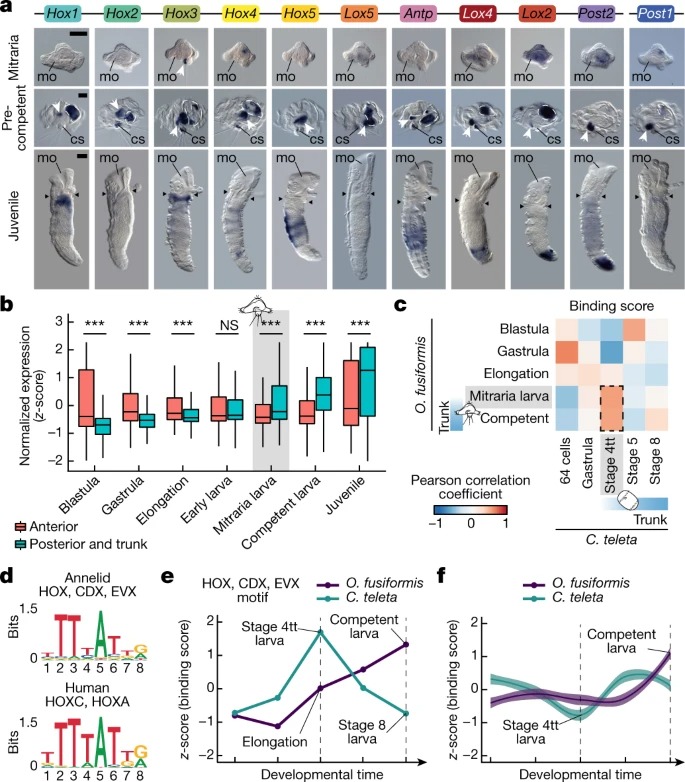
Fig. 3. 몸통 development은 O. fusiformis에서 pre-metamorphosis로 지연된다.
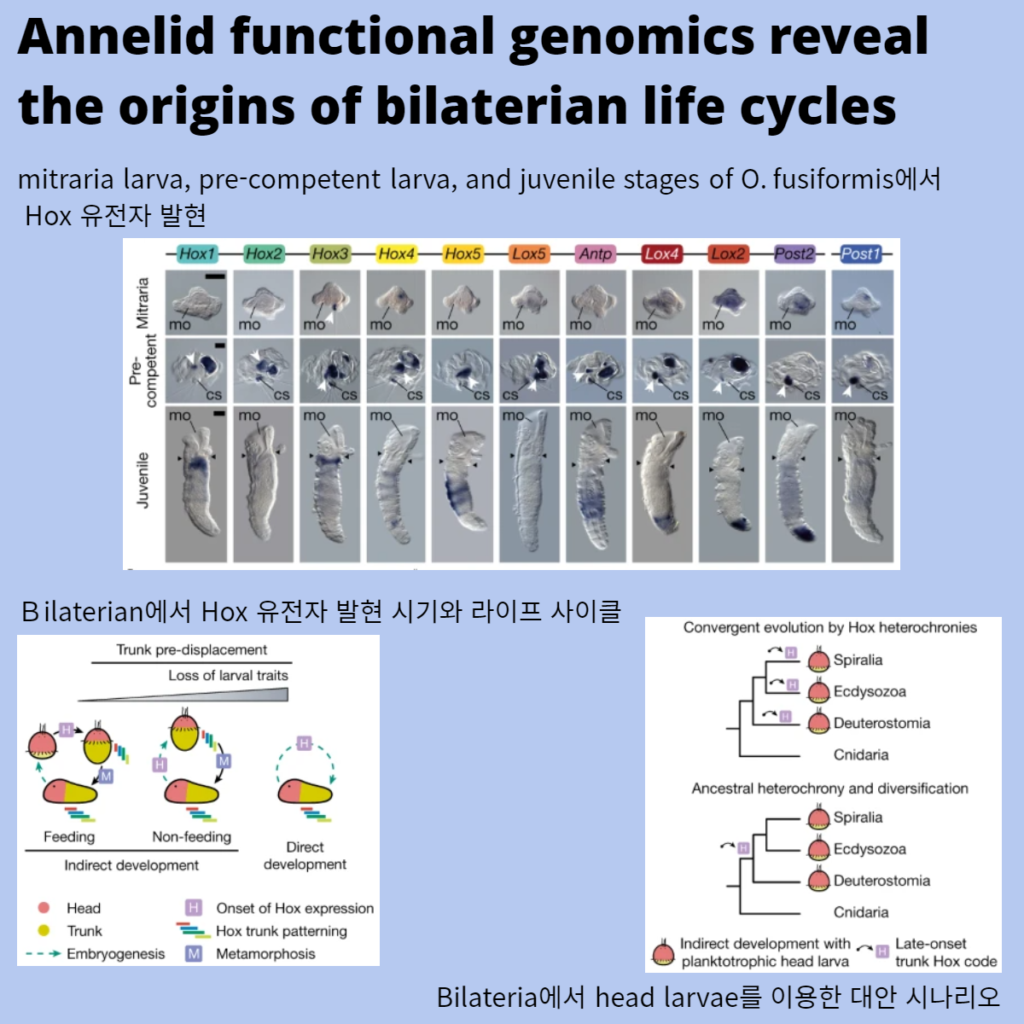
a, mitraria larva, pre-competent larva, and juvenile stages of O. fusiformis에서 whole-mount in situ hybridization에 의한 Hox 유전자 발현의 세 가지 독립적인 분석의 대표적인 이미지.
b, O. fusiformis 발달 중 해당 성체 조직에서 발현되는 anterior (n = 1,655)와 posterior and 몸통 gene (n = 407)의 Average expression dynamic.
c, transcription factor binding score (TFBS)의 Correlation matric.
d, annelid archetype (위)의 Sequence logo는 human homologue (아래)와 상당한 유사성을 보여준다.
e, O. fusiformis and C. teleta development 중 환형 HOX, CDX 및 EVX motif에 대한 TFBS dynamics.
f, Hox 클러스터의 피크에서 모든 motif의 평균 TFBS dynamics.
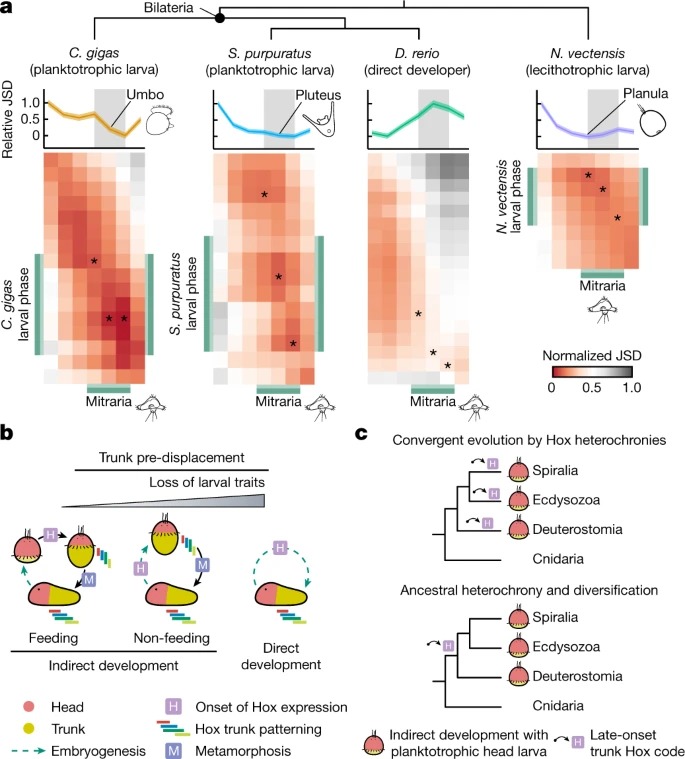
Fig. 4. Annelida와 Bilateria의 라이프 사이클의 진화.
a, O. fusiformis 와 Crassostrea gigas, Strongylocentrotus purpuratus, D. rerio and Nematostella vectensis 사이의 쌍방향 정규화된 Jensen–Shannon divergence (JSD)의 Heatmaps.
b, bilaterian에서 Hox 유전자 발현 시기와 세 가지 주요 유형의 라이프 사이클에 대한 도식.
c, Bilateria에서 head larvae를 이용한 maximal indirect development의 진화를 위한 대안 시나리오제안.
Top, head larvae는 (Hox 유전자 발현에서 볼 수 있듯이) 몸통 development를 반복적으로 pre-metamorphic stage로 이동시킴으로써 융합적으로 진화했다.
Bottom, 머리와 몸통의 발달은 ancestrally 시간적으로 분리되어 있었고, 이는 서로 다른 bilaterian 계통에서 head larvae의 진화를 촉진할 수 있었을 것이다.
Disscussion
우리의 연구는 Bilateria의 라이프 사이클 진화에 대한 관점을 제공한다. O. fusiformis의 플planktotrophic larva는 몸통 differentiation을 pre-metamorphic stage로 미루고 주로 ectodermal 영역에서 발달한다. 이것은 다른 먹이를 주는 환형 larvae (Extended Data Fig. 5f)에서 발생하며, 아마도 Chaetopteriformia에서 발생하며, 따라서 dult 몸통의 late differentiation은 환형 larvae의 조상적 특성일 수 있다(Extended Data Fig. 10b). 몸통 development를 post-larval stages로 늦추는 것은 Spiralia, Ecdysozoa and Deuterostomia의 계통발생학적으로 멀리 떨어진 분지에서도 발생하며, larvae는 일반적으로 head larvae라고 한다. 대조적으로, non-feeding larvae와 Annelida와 다른 bilaterian 분류군의 직접적인 developer들은 그들의 몸통을 anterior or head patterning하기 시작하는데, 이 패턴은 bilaterian에서 항상 gastrulation되기 전에 일어난다. 따라서, 몸통 development의 heterochrony는 동물의 다양한 라이프 사이클의 진화와 관련이 있으며, 이를 설명할 수 있다(Fig. 4b). 이것은 동물의 수명 주기의 기원을 설명하기 위해 이전에 제안된 메커니즘, 즉 larval-specific regulatory program에 성인 유전자의 co-option과 adult gene regulatory module의 독립적인 진화와 다르다.
Bilaterian head larvae는 몸통 differentiation와 Hox patterning을 지연시킴으로써 융합적으로 진화한 maximal indirect development의 진화와 관련된 lineage-specific innovation일 수 있다(Fig. 4c). larval molecular pattern의 유사성은 유사한 세포 유형과 larvae 기관을 개발하기 위해 독립적으로 협력된 ancient gene regulatory module을 반영할 것이다. 대안적으로, 몸통 differentiation와 Hox 발현의 post-embryonic onset은 Bilateria에 대한 가장 parsimonious ancestral state일 수 있다(Extended Data Fig. 10c,d and Supplementary Table 93). 이것은 larvae의 진화를 촉진할 수 있었고, larvae는 원래 발달을 위해 anterior genetic module을 공유할 것이다(Fig. 4c). 시나리오와 상관없이, 그리고 한계에도 불구하고, 우리의 데이터 세트는 bilaterian 라이프 사이클의 다양화를 위한 heterochronic change의 중요성을 강조한다. 이 데이터는 또한 Annelida와 심지어 Bilateria의 수명 주기 차이에 영향을 미칠 수 있는 후보 유전자와 regulatory motif의 감소된 세트를 발견한다. 앞으로 이들 유전자와 다른 유전자에 대한 비교 기능 연구는 유전자 발현과 조절의 시간적 변화가 동물 larvae와 성체의 진화를 어떻게 형성했는지 밝혀낼 것이다.