중세 독일 유대인의 게놈 전체 데이터에 따르면 아슈케나지 창립 사건은 14세기 이전에 일어났습니다.
Abstract
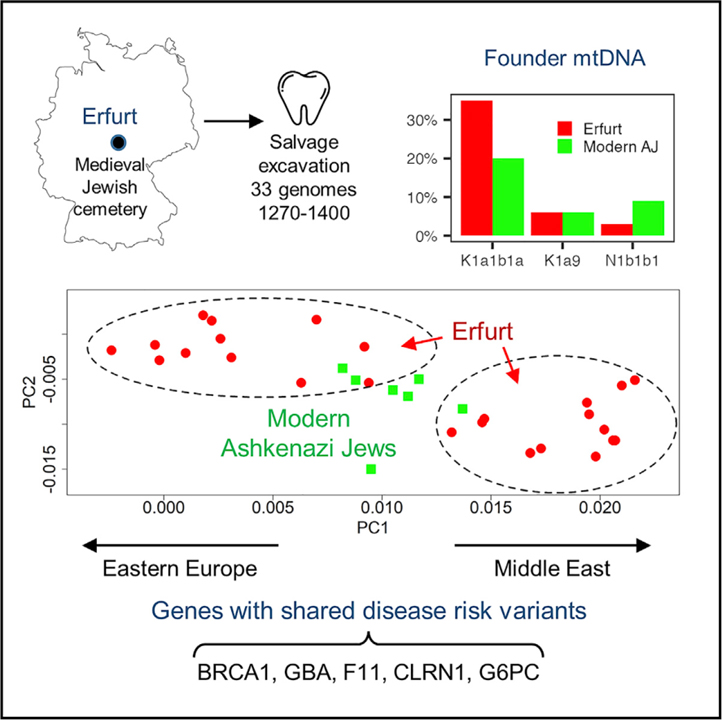
우리는 독일 에르푸르트의 중세 유대인 묘지에서 인양 발굴 후 얻은 14세기로 거슬러 올라가는 33명의 아슈케나지 유대인 (AJ)의 게놈 전체 데이터를 확인했습니다.
에르푸르트 개체는 현대 AJ와 유전적으로 유사하지만 현대 AJ보다 동유럽 관련 조상에서 더 많은 변이성을 보입니다. 에르푸르트 개체의 3분의 1은 현대 AJ에서 흔한 미토콘드리아 혈통을 가지고 있었고 8개는 오늘날 AJ에 영향을 미치는 것으로 알려진 병원성 변이체를 가지고 있었습니다. 높은 수준의 동형 접합성과 함께 이러한 관찰은 에르푸르트 커뮤니티가 이미 현대 AJ에 영향을 미치는 크기의 주요 감소를 경험했음을 보입니다. 에르푸르트 병목 현상은 더 심각하여 중세 AJ의 하부 구조를 암시합니다.
전반적으로, 우리의 결과는 AJ 창립 사건과 조상의 주요 출처 획득이 14세기 이전으로 거슬러 올라가고 현대 AJ에 더 이상 존재하지 않는 중세 후기 유전적 이질성을 강조한다는 것을 시사합니다.
Figure
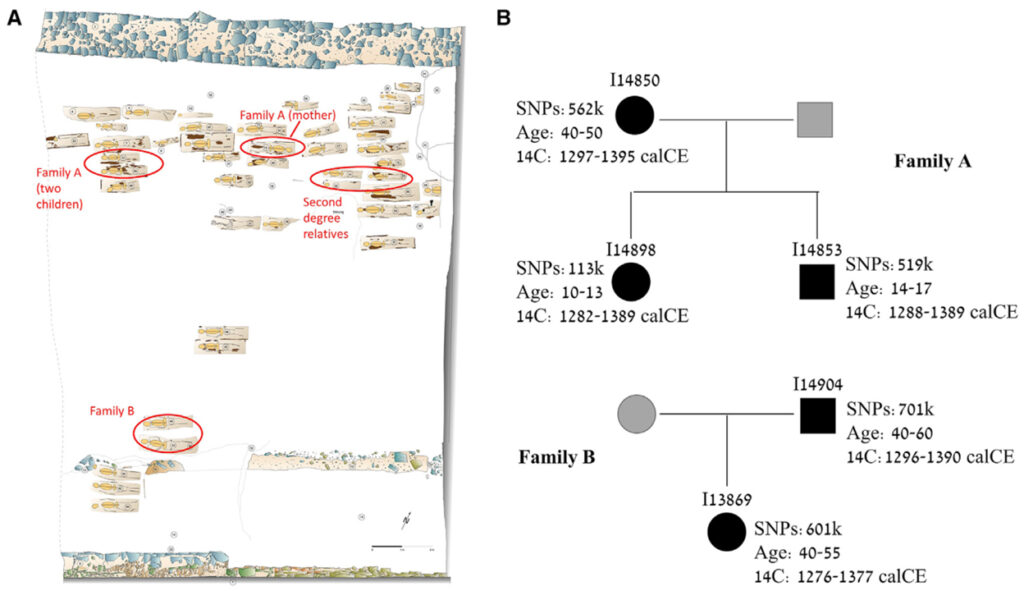
[Figure 1] 에르푸르트의 중세 유대인 묘지 (A) 묘지의 배치. 내부 성벽과 외부 성벽은 각각 지도의 하단과 상단에 있습니다. 가족 구성원은 빨간색 타원으로 표시됩니다. (B) 1촌 관계에 따라 확인된 두 가족의 혈통. 검정색 기호는 DNA를 가진 개인을 나타냅니다. 회색 기호는 유추된 가족 구성원을 나타냅니다. 원: 암컷; 사각형: 수컷. 각 개인에 대해 ID, 유전자형 SNP 수, 예상 사망 연령 및 14C 날짜 (95.4% 확률 간격)를 표시합니다.
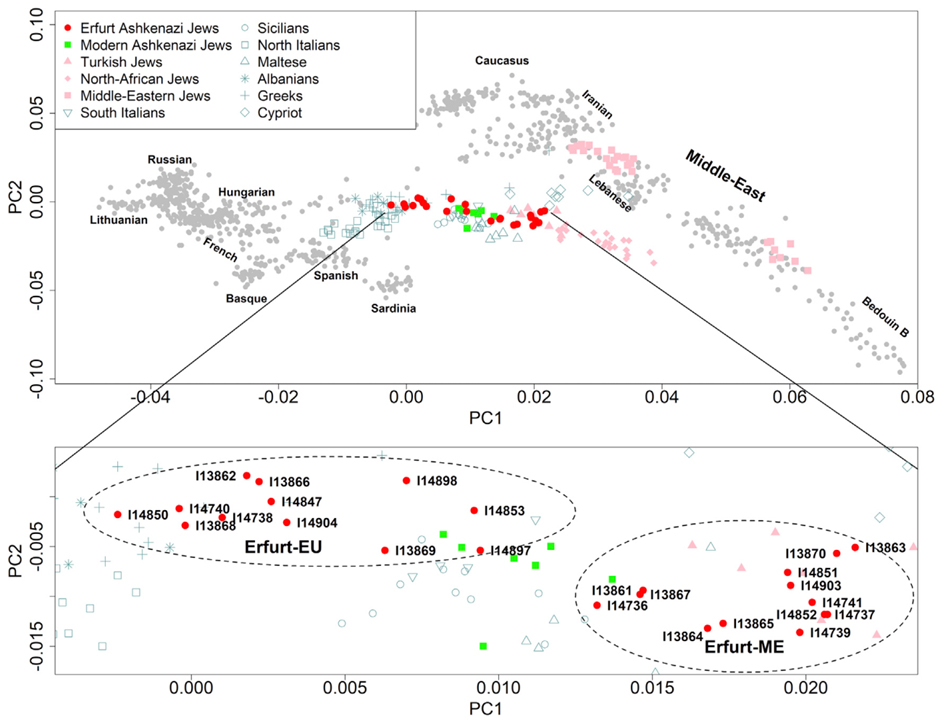
[Figure 2] 주성분 분석 우리는 서부 유라시아 인구를 사용하여 주성분 (PC)을 배웠고 에르푸르트 개체 (빨간색 원)를 추론된 축에 투영했습니다. 현대의 아슈케나지 유대인 (녹색 사각형), 아슈케나지 출신이 아닌 유대인 (분홍색 모양), 지중해 인구 (청록색 모양)가 강조 표시되어 있습니다. 삽입된 부분은 AJ 개인이 포함된 지역을 확대합니다.
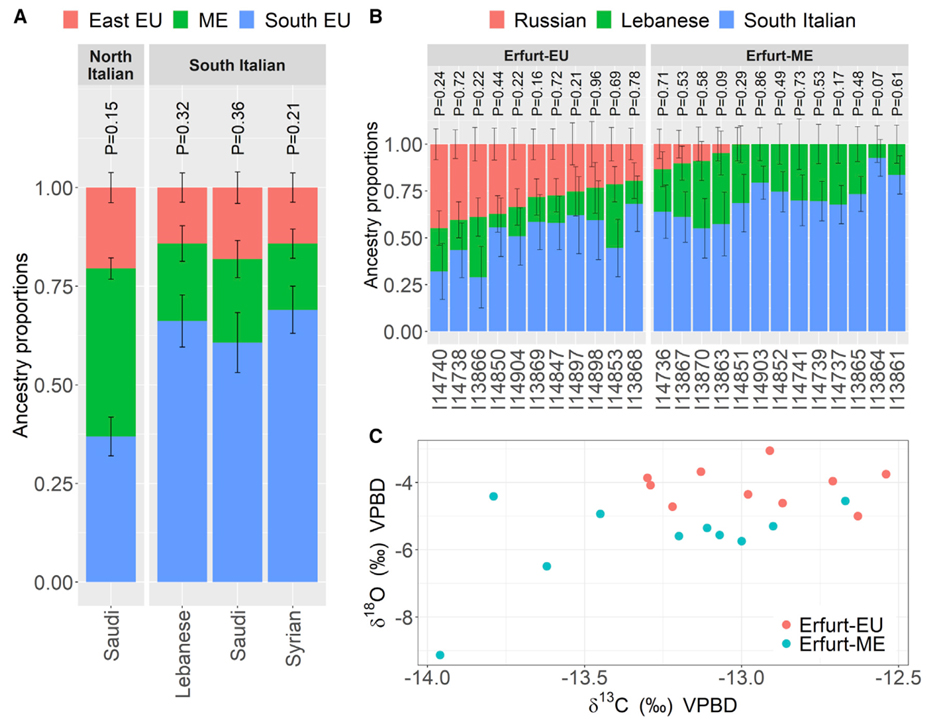
[Figure 3] 에르푸르트 아슈케나지 유대인 조상 모델 (A) 에르푸르트 유대인의 조상에 대한 각 qpAdm 모델에는 중동, 남부 유럽 및 동유럽 (러시아인) 소스가 포함됩니다. 남유럽 출처는 각 패널의 상단에 표시된 대로 남이탈리아인 또는 북이탈리아인이었습니다. 중동 소스는 x 축 레이블에 표시됩니다. 주요 분석 및 견고성 테스트에서 qpAdm p 값 >0.05인 모델만 표시됩니다. 오차 막대는 각 방향에서 하나의 표준 오차를 나타냅니다. qpAdm p 값은 각 모델 위에 표시됩니다. (B) ID로 표시된 단일 에르푸르트 개인의 조상. 러시아, 레바논 및 남부 이탈리아 소스와 함께 qpAdm을 사용했습니다. 개인은 에르푸르트 하위 그룹 (EU/ME)으로 분류됩니다. qpAdm p 값은 각 개인에 대해 표시됩니다. 적용 범위가 낮은 개인 (<50k SNP) 및 이러한 소스를 사용하여 모델링할 수 없는 추가 개인 (p < 0.05)에 대해서는 결과가 표시되지 않습니다. (C) >200k SNP를 갖는 20개 에르푸르트 개체의 하위 집합에 대한 d13Cenamel 및 d18Oenamel 안정 동위원소 비율의 플롯. 에르푸르트 하위 그룹 소속 (EU/ME)은 색상으로 구분됩니다.
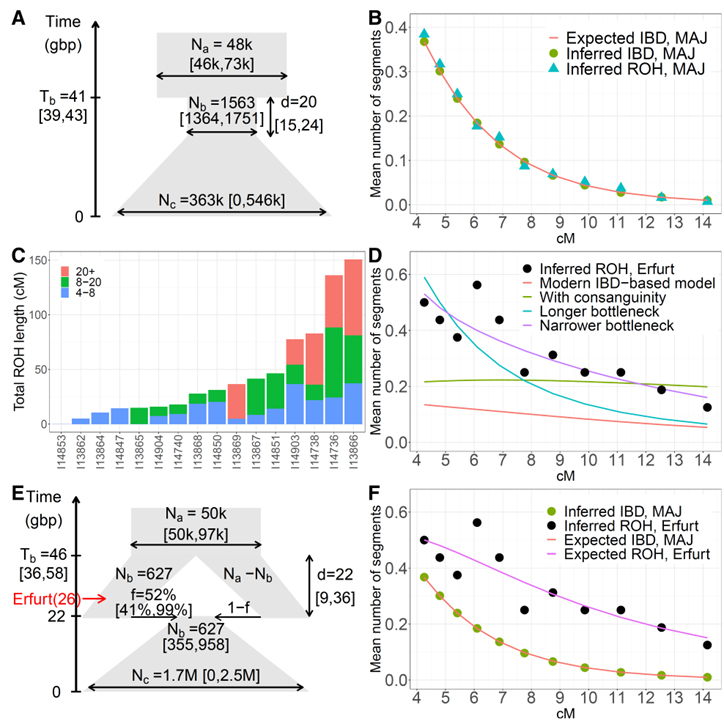
[Figure 4] 고대 및 현대 일배체형에 기반한 AJ 인구통계학적 이력 모델 (A) 현대 IBD 공유를 기반으로 추론된 AJ의 인구 통계학적 이력에 대한 단일 인구 모델(표 S5, 모델 [A]). 다이어그램에서 y축은 현재 이전 세대의 시간 (gbp)을 나타내며 너비는 유효 인구 크기에 (도식적으로) 비례합니다. 95% 신뢰 구간 (CI)은 부트스트래핑을 사용하여 계산되었으며 각 매개변수 근처에 표시됩니다. (B) 최신 AJ 대 세그먼트 길이 (4에서 15cM 사이의 11개 빈)에서 IBD 및 ROH 세그먼트의 평균 수 (반수체 상염색체 게놈 쌍당). 각 기호 (IBD의 경우 원, ROH의 경우 삼각형)는 해당 빈의 중앙에 배치됩니다. 빨간색 선은 (A)의 인구 통계학적 모델을 기반으로 빈당 예상되는 세그먼트 수를 보여줍니다. (C) >400k SNP를 가진 16명의 EAJ 개체에서 ROH 세그먼트의 총 길이. 막대는 길이가 다른 세그먼트의 기여도에 비례하여 색상이 지정됩니다. (D) ROH는 EAJ (원)로 계산하고 다양한 모델 (선)을 기반으로 예상되는 수입니다. 최신 IBD를 사용하여 추론된 모델은 빨간색입니다. 동일한 모델이지만 EAJ에서 혈연관계를 허용하는 것은 녹색입니다. 두 모델 모두 짧은 ROH 세그먼트에 적합하지 않습니다. (A)와 유사하지만 더 좁거나 더 긴 병목이 있는 모델은 각각 보라색과 청록색입니다. (E) MAJ의 IBD와 EAJ의 ROH를 사용하여 공동으로 추론한 두 모집단 모델. 모델에 따르면, 조상 개체군은 Tb = 46세대 전에 유효 크기 Nb = 627 (빨간색으로 표시된 에르푸르트를 나타냄)의 한 개체군과 Na Nb 크기의 다른 개체군으로 분할했습니다. 병목 현상이 끝나면 두 모집단은 각각 f = 52% 및 1 f 비율로 병합되고 단일 모집단 모델에서와 같이 기하급수적으로 확장됩니다. 에르푸르트 인구의 샘플링 시간은 26세대 전으로 표시됩니다 (세대당 25년으로 가정). (F) MAJ의 IBD 세그먼트 수(녹색 원, [B]와 동일한 데이터) 및 EAJ의 ROH (검은색 원, [D]와 동일한 데이터) 및 두 모집단을 기반으로 하는 기대치 (선) (E)의 모델 (MAJ, 빨간색, EAJ, 분홍색).
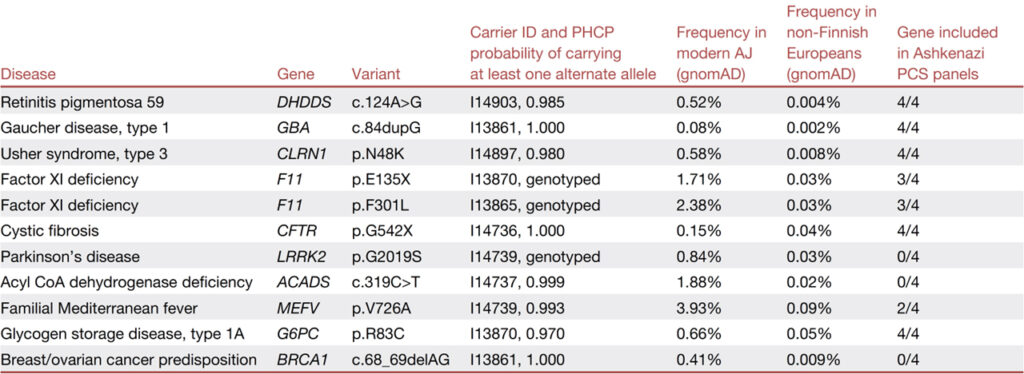
[Table 1] 에르푸르트에서 검출된 신뢰도가 높은 AJ 농축 병원성 변이체 11개 변이 각각에 대해 HGVS (Human Genome Variation Society) 명명법으로 질병, 유전자 및 변이를 표시합니다. c.68_69delAG BRCA1 변형은 185delAG라고도 합니다. 풍부한 SNP를 포함하는 서열에서 검출된 변이체는 “유전자형”으로 지정됩니다. 귀속 변이체의 경우 PHCP에서 적어도 하나의 대체 대립유전자를 갖는 한계 사후 확률을 제공합니다. 추가로 캐리어 ID, MAJ 및 비핀란드 유럽인 (gnomAD)의 대립유전자 빈도, 유전자가 포함된 아슈케나지 특정 임신 전 검사 (PCS) 패널의 수 (4개 중)를 제공합니다.
Disscussion
이 연구는 역사적인 AJ 개인의 게놈 전체 데이터를 분석하여 초기 AJ 기원에 대한 이해를 개선합니다. EAJ의 조상은 PCA, ADMIXTURE, qpWave 분석에서 입증된 바와 같이 MAJ의 조상과 밀접한 관련이 있으며, 이는 지난 700년 정도의 시간 동안 AJ 조상의 높은 수준의 연속성을 시사합니다. 그러나 EAJ 개인은 MAJ보다 더 많은 변이 조상을 가지고 있었고 소수의 동유럽 관련 조상 구성 요소의 존재로 계층화되었습니다. 여러 증거에 따르면 EAJ 인구는 이미 MAJ와 공유되는 “병목 현상”을 경험했습니다. 즉, 아슈케나지 설립자 mtDNA 혈통의 높은 빈도, AJ가 풍부한 병원성 변이체 및 기타 대립 유전자의 존재, 장기간의 동형 접합성입니다. 이전 연구와 일치하여 우리는 AJ 확장의 시작을 약 20-25세대 전에 시작했습니다.
우리의 고대 DNA 데이터를 통해 AJ의 역사에서 현대의 유전적 변이에서 감지할 수 없었던 패턴을 식별할 수 있었습니다. 특히, 우리의 결과는 AJ 인구가 오늘날보다 중세 시대에 더 구조화되었음을 시사합니다. 에르푸르트 내에서 한 그룹은 동유럽 관련 조상이 많았고 다른 그룹은 서유럽 출신의 MAJ와 현대 세파르디 유대인에 가까운 조상을 가졌습니다. 두 그룹은 또한 에나멜 δ18O의 뚜렷한 수준을 가졌습니다. 우리의 결과는 에르푸르트 내에서 더 미세한 구분도 배제할 수 없으며 중세 AJ는 에르푸르트 너머에 추가로 구조화되었을 수 있습니다. 대조적으로 오늘날의 AJ는 현저하게 동질적입니다. 이것은 조상의 전반적인 출처가 중세와 현대 AJ 사이에서 유사하게 유지되었지만 중세 이후의 내혼 및 AJ 내 혼합이 AJ 유전자 풀의 균질화에 기여했음을 시사합니다.
우리는 중세 후기에 AJ 인구가 서쪽/동쪽 축을 따라 언어적으로나 문화적으로 나뉘었음을 암시하는 비유전적 데이터를 기반으로 두 에르푸르트 하위 그룹의 정체성을 추측할 수 있습니다. 에르푸르트-ME에 해당할 수 있는 서부 AJ 그룹은 라인란트의 초기 AJ 정착민의 후손을 대표했을 가능성이 높습니다. 한 소식통은 에르푸르트가 두 그룹 사이의 경계에 있다고 명시적으로 언급하며, 14세기에 에르푸르트의 이름은 서부 및 동부 AJ의 전형이었습니다. 이러한 점은 동부에서 두 번째 에르푸르트 공동체로 이주한 증거와 함께 두 유전 그룹에 속하는 개인이 에르푸르트에 존재한 이유를 설명할 수 있습니다. 에르푸르트의 유전적 다양성 증가는 EAJ가 MAJ보다 더 심각한 병목 현상을 경험했다는 우리의 추론과 모순되지 않습니다.